耳石的主要功能為聽覺及感覺平衡作用。除了這些生理功能之外,最特別的地方是耳石扮演著計時器和環境紀錄器的功能。常言道:走過的必留下痕跡。魚類何時游過何種環境,會在其耳石中留下蛛絲馬跡。耳石的研究是近20年來最吸引魚類生態學家的課題之一。透過耳石日周輪或年輪以及微化學的分析可以再現魚類過去的洄游環境史。
1.5.1 耳石是什麼玩意(What is an otolith?)
耳石(Otolith, ear stone)是生物礦化作用所形成的碳酸鈣結晶。魚類的內耳迷路(Vestibular apparatus)中有三對耳石:分別為矢狀石(Sagitta)、礫石(Lapillus)以及星狀石(Asteriscus)。三對耳石的大小不一,矢狀石隨著魚體的成長同步增大,其他兩對耳石的相對成長速度緩慢(圖1.5.1)。因矢狀石體積
比較比其他兩對大,
方便處理而且比較能代表魚體的成長,一般都用它來當做研究
魚類成長和洄游環境的
媒介代言者。
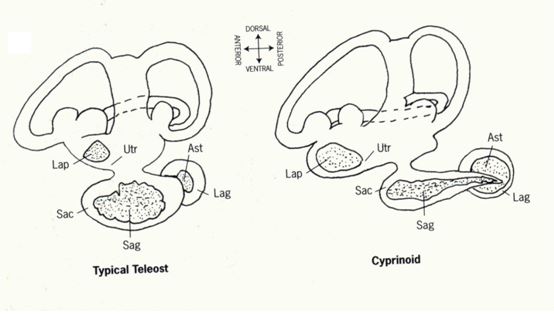
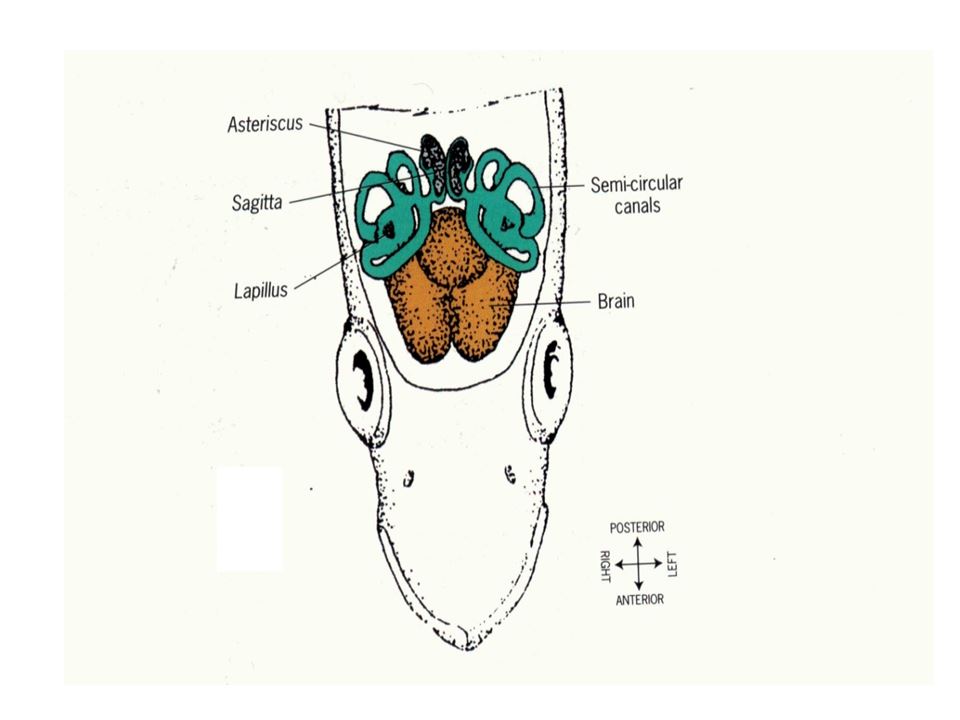
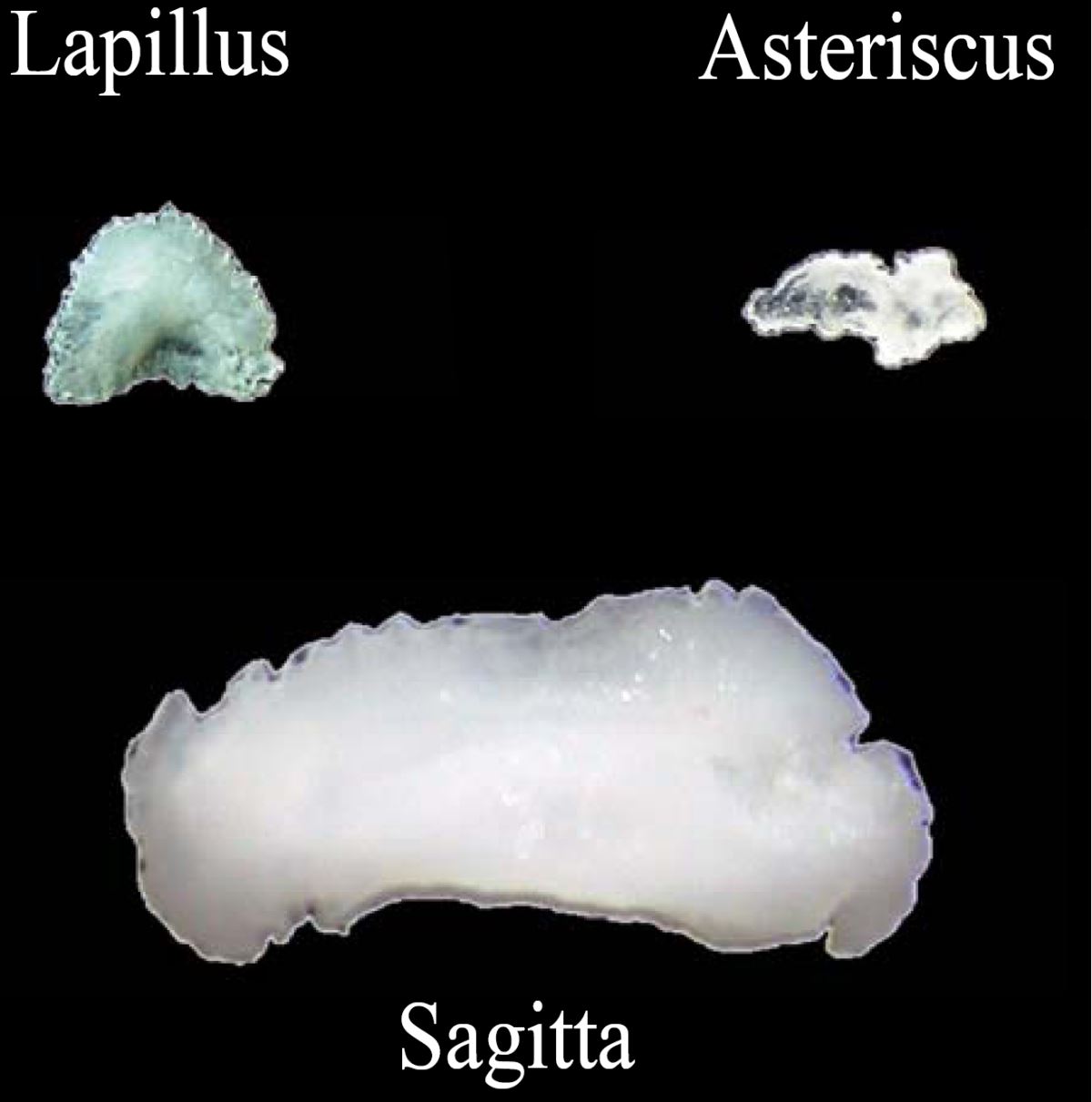
圖1.5.1魚類的三對耳石:矢狀石(Sagitta, Sag)、礫石(Lapillus, Lap)以及星狀石(Asteriscus, Ast)。(a) 典型真骨魚類與鯉科魚類的內耳迷路之比較(Lowenstein 1971)。 (b)真骨魚類內耳迷路的背面觀(Secor
et al. 1992) 。(c) 三對耳石的相對大小(photo: 許智傑)
1.5.2 會寫日記的魚類(Fishes can write diary)
耳石的沉積速率,因魚類成長受日夜光週期的影響而產生日周輪,受季節性變化的影響而產生年輪。早在100多年前德國科學家Reibisch就知道利用耳石上的年輪來測定比目魚(
Pleuronectes platessa)的年齡。耳石日周輪是美國耶魯大學的地質學教授潘尼拉博士(Pannella,1971,Science 173: 1124-27),在一次偶然的機會,從沙灘的沉積物中發現的,從此魚類的定齡精準度便由年的尺度提升到日的層次,進而揭開了魚類很多未知的初期生活史秘密。因為日周輪的發現,被稱為會寫日記的魚類。
透過電子顯微鏡的解析,發現耳石日周輪包括一個成長帶(incremental zone)和一個不連續帶(discontinuous zone)(Tzeng 1990,Tzeng and Yu 1988),成長帶是魚類白天生長旺盛時形成的,不連續則是清晨空腹時形成的。兩個帶所形成的生物時鐘時間大約24小時,也就是一天形成一輪。所以由日周輪數目可推算魚類的生日和日齡(圖1.5.2)。
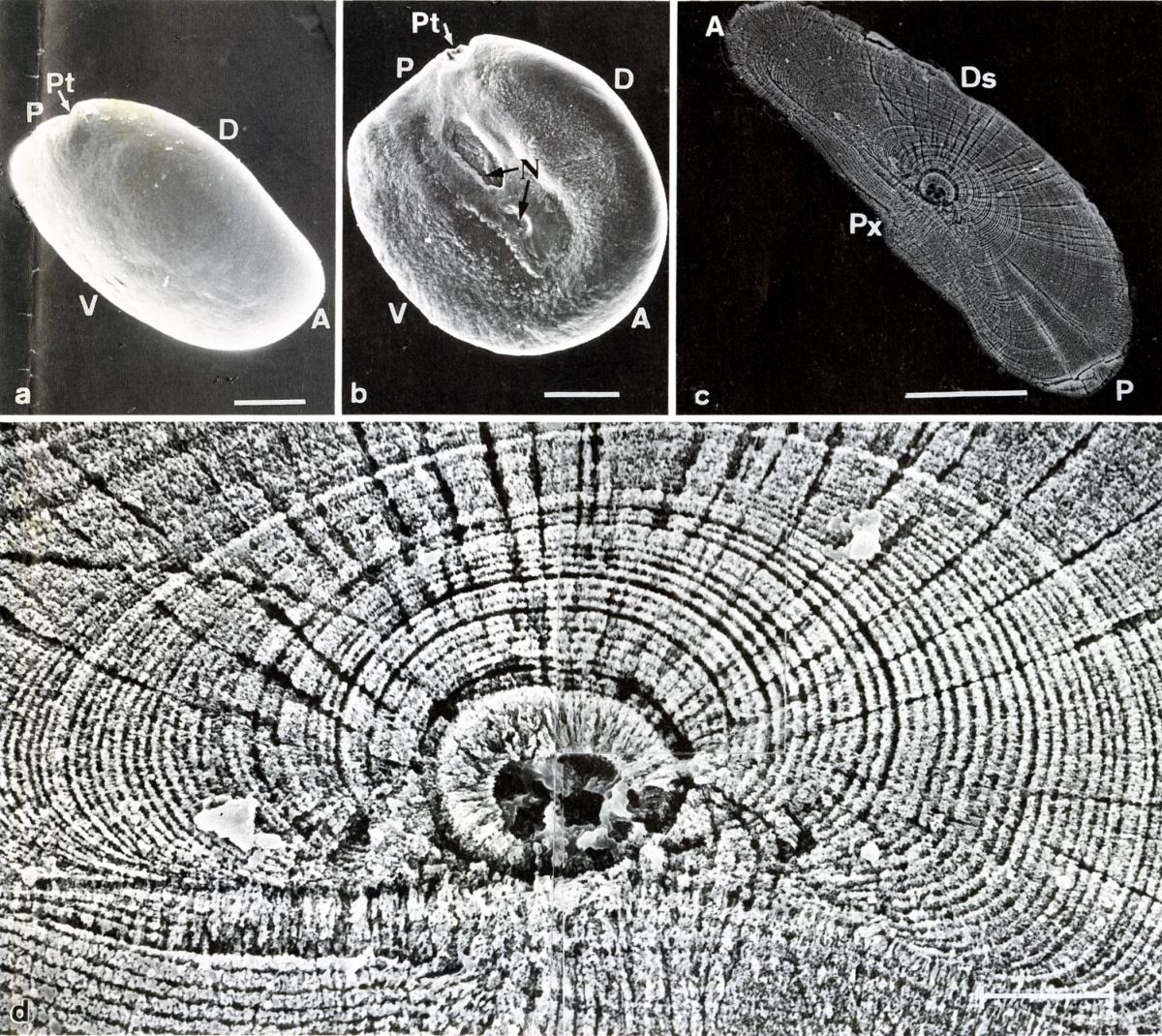
圖1.5.2 這是臺灣第一張耳石日周輪的掃描式電子顯微鏡照片。(上圖)分別為日本鰻鰻線耳石的外側觀(a)、內側觀(b)以及耳石的縱切面(c), (下圖)(c)圖日周輪的放大(Tzeng 1990)
1.5.3 耳石有如羅塞達石碑也有點像電腦的光碟片(Otolith like Rosetta stone and CD-ROM)
有人把耳石比喻為1799年在尼羅河所發現的羅塞達石碑(Rosetta stone)(圖1.5.3)。這個石碑同時以羅馬文字,埃及文字及古埃及的象形文字記載著西元前196年埃及國王托勒密五世(Ptolemy V)的詔書。因為這塊石碑的出土,古埃及的象形文字得以破解。耳石日周輪上的刻痕記載著
鰻魚
類的每一個發育階段
和的生活史細節,透過日周輪的解讀不是也能夠破解鰻魚
過去的生活史秘密嗎?(Tzeng 1990, 2003, Cheng and Tzeng 1996, Wang and Tzeng 1998, 2000)。

圖1.5.3(圖左)羅塞達石碑,(圖右)玻璃鰻耳石日周輪與鰻魚發育階段的對應關係。P:核心, HC:孵化輪, FFC:第一次攝食輪, MC: 變態輪, EC:鰻線輪
鰻魚耳石的核心部分是在卵黃囊仔魚期時所形成的,而卵黃是來自母親,因此這一部份是母源效應,其反應的是親代的生活狀況。第一個日周輪是仔魚開始吸收外界物質後所形成的。之後每一天形成一輪,因此由日周輪數可以推算期孵化日,日齡及日成長率等(Tzeng 1990,Tzeng & Tsai 1992,Wang and Tzeng 1998,2000,Shiao
et al. 2001,2002)。
從某個角度來看,耳石更像電腦的光碟片(CD-ROM)(圖1.5.4)。圖中的耳石,是1995年8月22日台灣水產試驗所水試一號在菲律賓東邊的海域(14°00’N, 137°30’E)採集到的一尾從產卵場順著洋流漂過來的柳葉鰻,耳石經研磨到核心、EDTA腐蝕、電顯放大2000倍的照片。其形狀和同心圓的日周輪有些像電腦的光碟片。經解讀後,得知耳石有45圈日周輪,換算回去的誕生日期為7月3日,相當於農曆的新月期間。換言之,日本鰻是在夏天的新月夜間產卵(Liao
et al. 1996)。
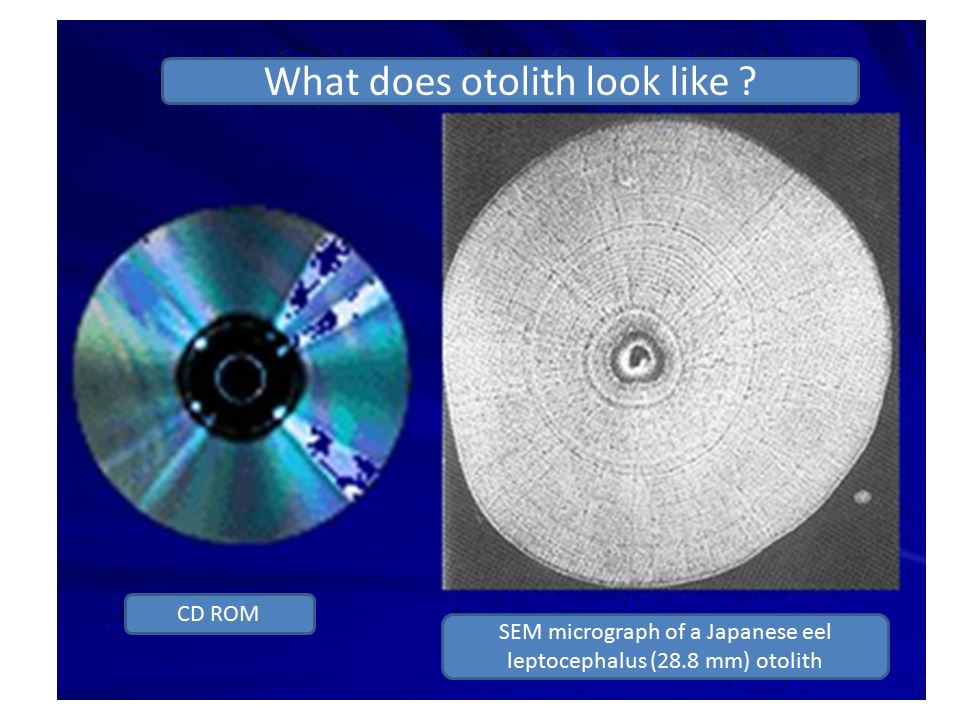
圖1.5.4(左圖)電腦的光碟片,(右圖)柳葉鰻(28.8um)的耳石電顯照片
1.5.4 耳石不但可以知道魚類的年齡而且可以知道魚類的日齡(Otolith can determine the age of fish in year but also in daily)
魚類的年齡,可利用耳石上的年輪來推定。因年輪是魚類的春夏秋冬四季成長速度不同而形成的。尤其是溫帶地區四季分明,魚類一年形成一輪的規則性非常明顯。例如生長在北溫帶
加拿大新斯科島(No
va Scotia)的美洲鰻,因冬季水溫降到零度以下而
停止生長,其耳石上的
年輪非常明顯(圖1.5.5)。可是生長在亞熱臺灣的日本鰻,冬季水溫不會很低,年輪就不會那麼明顯。由此可見,耳石不是告訴我們魚類的年齡,而且可以知道魚類的成長狀況和環境的關係(詳曾萬年教授編著(2012) 鰻魚生活史及保育論文集
(上、下冊)。國立臺灣大學圖書館,『臺大人』文庫)。

 圖1.5.5 加拿大美洲鰻耳石縱切面的光學顯微鏡照片。耳石的明帶是春夏秋季鰻魚生長速度快時形成的,暗帶(年輪)是冬季生長停滯時形成的(上圖
圖1.5.5 加拿大美洲鰻耳石縱切面的光學顯微鏡照片。耳石的明帶是春夏秋季鰻魚生長速度快時形成的,暗帶(年輪)是冬季生長停滯時形成的(上圖:
年齡約12歲,下圖:18歲)
魚類的耳石除了年輪記號之外,如果將耳石用電子顯微鏡放大2000倍的話,還可以看到比用光學顯微鏡放大所看到的年輪更更細微的日周輪,因日周輪是受日夜明暗周期的生理時鐘的調控,每天形成一輪。日周輪數目相當於魚類的日齡,利用耳石上的日周輪可推算鰻線到達河口域時候的日齡及其生日。圖1.5.6是日本鰻鰻線和美洲鰻鰻線的日日周輪。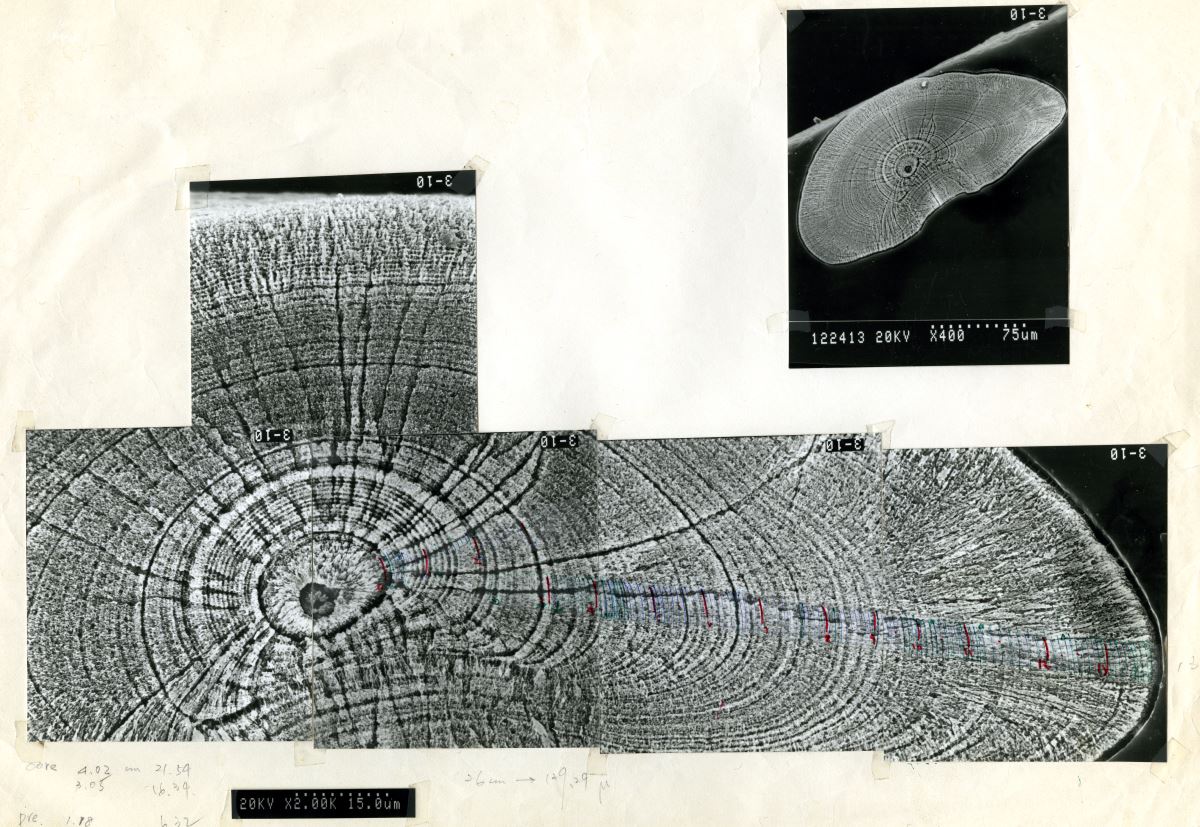
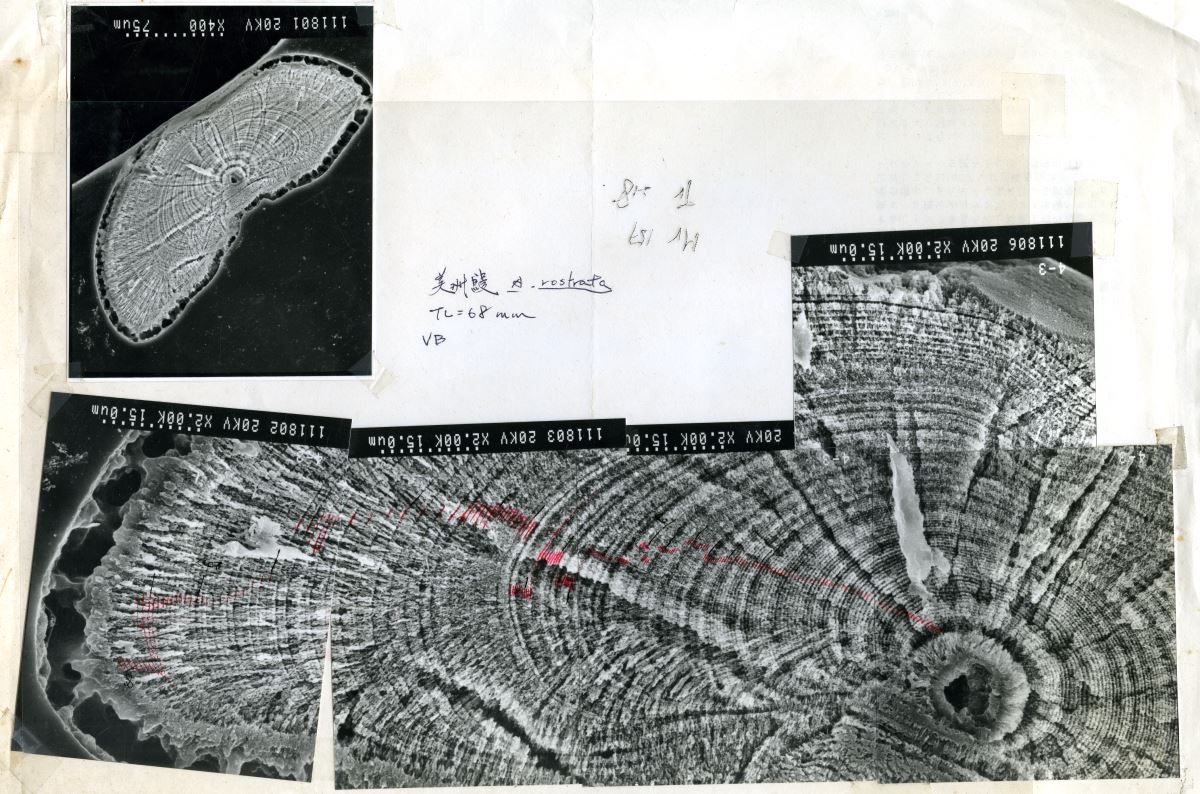 圖1.5.6 放大2000倍的鰻線耳石日周輪電子顯微鏡照片。明帶市白天形成的,暗帶則是夜間形成的 (上圖
圖1.5.6 放大2000倍的鰻線耳石日周輪電子顯微鏡照片。明帶市白天形成的,暗帶則是夜間形成的 (上圖:
日本鰻,下圖:
美洲鰻 )
1.5.5 耳石的微量元素組成可重建魚類的洄游環境史(Otolith elemental conposition can reconstruct the migratory environmental history of the fish)
耳石中的化學元素大約有31種左右(Panfili et al. 2002, Fig.VII.3)。耳石隨著魚體的成長而增大,同時沉積魚類洄游環境中的化學元素。當我們利用電子微探儀(EPMA)或雷射藕合電漿質譜儀(LA-ICPMS),在耳石的切面上從核心至耳石邊緣掃描時,可以測量這些元素隨魚體成長過程的變化,用來重現魚類從出生到被捕獲為止的生活史及所經歷的環境。以1994年在臺灣北部淡水河捕獲的玻璃鰻的耳石上的日周輪和Sr/Ca為例,可以知道這隻玻璃鰻從出生到臺灣北部淡水河的每一發育階段的持續天數和洄游環境(圖1.5.
75a,b)。這是臺灣第一張耳石Sr/Ca比的測量技術圖。
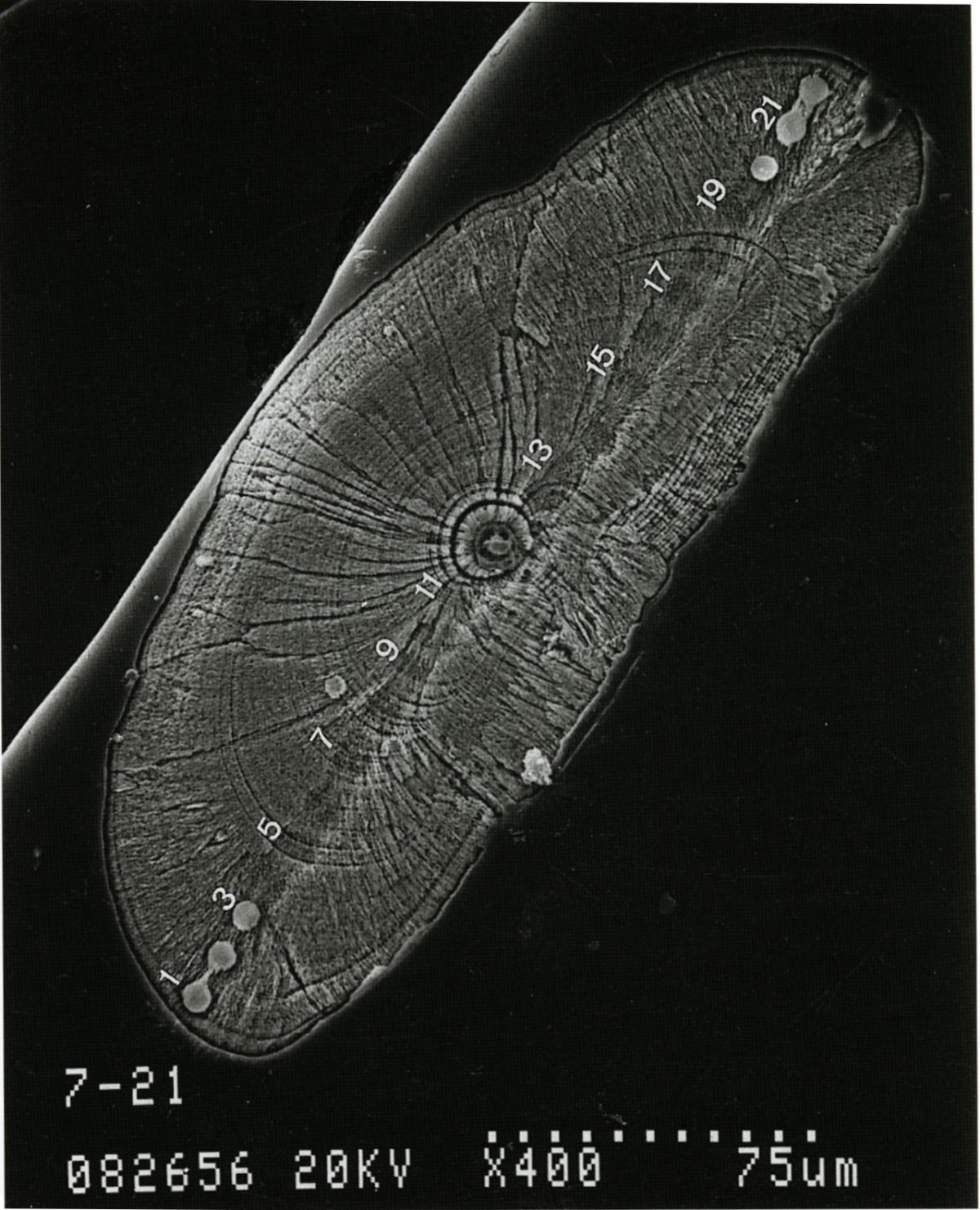
圖1.5.
7a電子微探儀(EPMA)測量耳石Sr/Ca比時的掃描點(Tzeng and Tsai 1994)
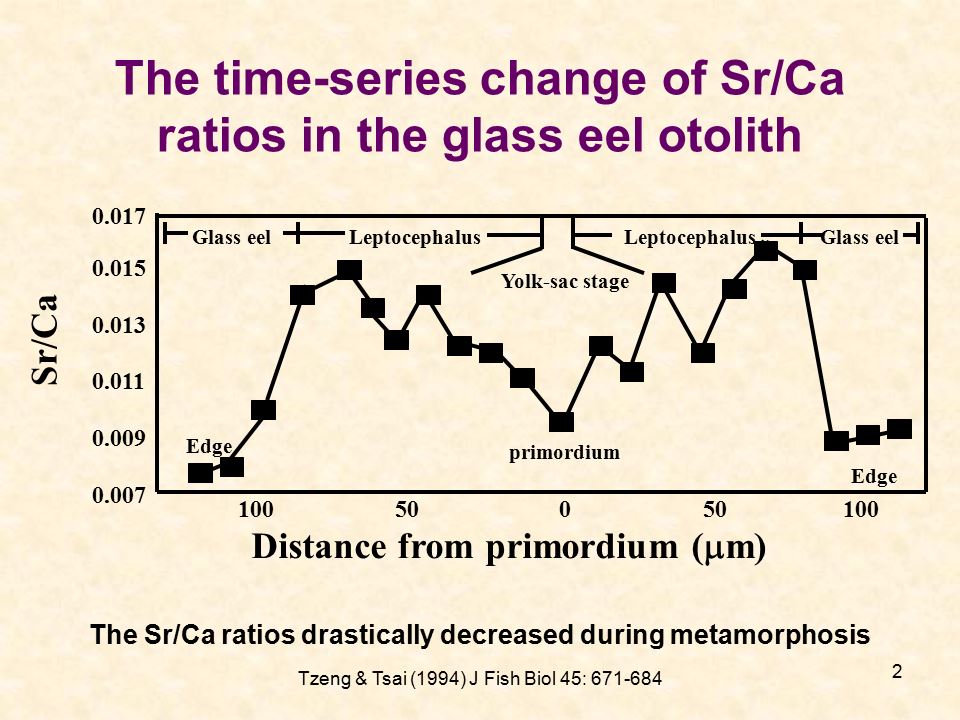
圖1.5.
7b玻璃鰻耳石Sr/Ca比隨發育階段的變化。
鍶(Sr)是研究耳石微化學最常用的元素,在化學週期表中與鈣(Ca)同屬鹼土族元素,最容易取代耳石中的主要成份(碳酸鈣)中的鈣,而沉積在耳石中。海水中的鍶濃度是淡水環境中的100倍,因此從耳石中的Sr/Ca比的時間變化,便可以推測魚類在海水與淡水之間的洄游履歷(Tzeng and Tsai 1994; Tzeng 1995; Tzeng
et al. 2002, 2003)。
1.5.
5從耳石所看到的鰻魚從產卵場到陸地河川的洄游現象(The larval migration from ocean to estuary as revealed by otolith microstructure )
日本鰻柳葉鰻和玻璃鰻在海上的分布
,大體上以黑潮為界
。,黑潮的外洋側是柳葉鰻,黑潮
的陸地側是玻璃鰻(Tzeng 2003)(圖1.5.
86右圖)。
河口域採集到的鰻線耳石上的日周輪,也透露一些柳葉鰻漂流過程的訊息
(圖1.5.6左圖)。從台灣(T、S)、中國大陸(M、C and Y)及日本(I)等地採集的鰻線,其耳石日齡呈現由南往北增加的趨勢。鰻魚出生後從產卵場漂到東北亞國家需要5~6個月時間,愈北鰻線的日齡越大
(表二)(Cheng and Tzeng 1996)。
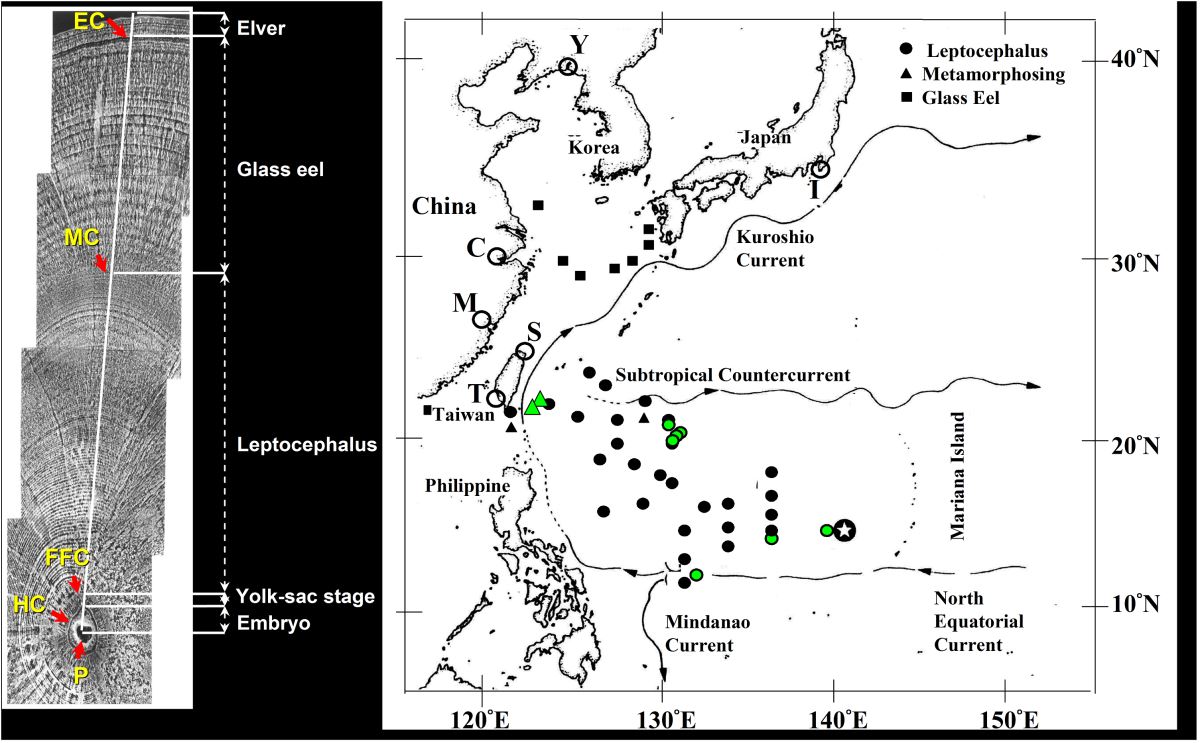
圖1.5.8 (右圖)日本鰻柳葉鰻(Leptocephalus)、變態中的柳葉鰻(Metamorphosing)、玻璃鰻(Glass eel)等發育階段在海上的分布以及河口域鰻線的採集點(T, S, M, C, Y, I)。星星記號: 產卵場位置,綠色點:臺灣的海上調查點,黑色點: 日本的海上調查點。(左圖) 日本鰻鰻線耳石上的日周輪及其發育階段
日本鰻從產卵場洄游到台灣河川入海口的洄游歷程及時間全都記錄在耳石上(Cheng and Tzeng 1996)。柳葉鰻順著北赤道洋流及黑潮向西北方向移動,大約經過4-5個月的海上漂游後抵達大陸棚區,然後變態成流線型且透明的玻璃鰻。再經過一個多月之後,來到河川入海口,碰到淡水之後身上出現黑色素,稱之為鰻線(elver)。
柳葉鰻的變態日齡(Tm)和 柳葉鰻變態之後至玻璃鰻來到河口的時間(Tr-m)是決定玻璃鰻進入那一個國家的關鍵因素。如果把Tm拿來做採集點間的比較,發現日本(I)與台灣(TS2)的平均Tm相差21天(表二),若與黑潮的流速(96km/天)相乘,得到的距離為2016公里,相當於台灣到日本的黑潮流路的距離(表二)。換句話說,早變態的柳葉鰻,其玻璃鰻就進入台灣,晚變態者進入日本(Cheng and Tzeng 1996)。
表二、柳葉鰻變態日齡(Tm) 和 柳葉鰻變態變態之後至玻璃鰻來到河口的時間(Tr-m)之地區間比較
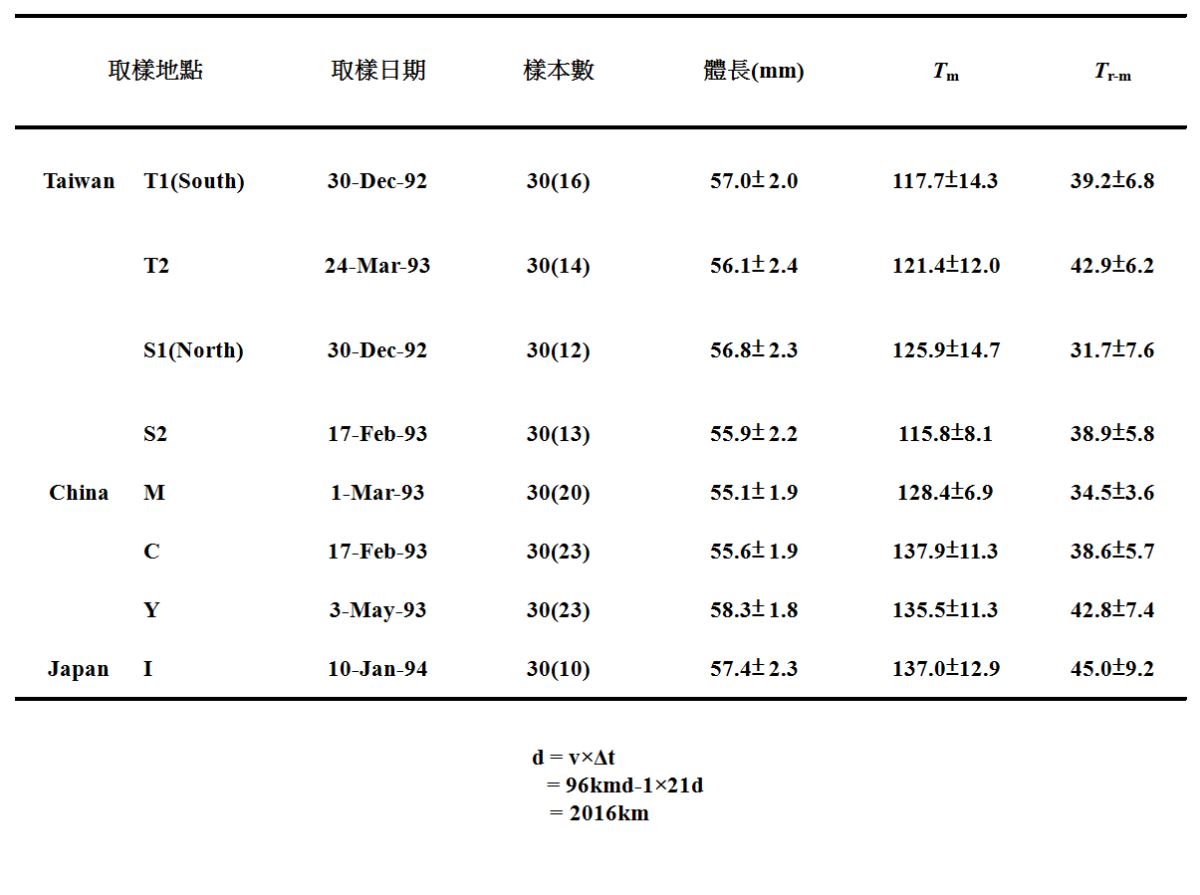
日本鰻誕生之後的向岸洄游的第二階段是柳葉鰻變態成玻璃鰻之後,脫離黑潮進入沿岸域的過程。如果我們把耳石上從變態到河口的這一段日周輪(Tr-m)做一個統計,會發現變態之後來到河口的時間需要一個月到一個半月(表二)。以台灣為例Tr-m是由北(S1,S2)往南(T1,T2)增加,顯示玻璃鰻可能是順著中國大陸沿岸流南下的,這個現象與台灣沿岸的日本鰻鰻線的漁獲量分佈一致(圖1.3.14)。
1.5.6耳石微化學的國際合作研究(International collaborative study on otolith microchemistry)
瑞典是第一個與台灣合作,進行歐洲鰻耳石微化學研究的歐洲國家。故事敘述著一隻歐洲鰻,從大西洋百慕達神祕三角洲附近的藻海順著海流漂泊,最後來到瑞典西海岸,總共在海中生活了9年,又被瑞典漁民把牠移植到東部Angen的淡水湖生活了12年,最後被送入人類的五臟廟。這些流浪過程的點點滴滴,都清清楚楚地寫在這隻歐洲鰻耳石中的年輪及Sr/Ca比的時間變化上(詳Tzeng
et al. 1997 的 Fig. 3c, 或Tzeng
et al. 1999 Fig. 1c, d)。換言之,耳石中的自然標記,年輪及Sr/Ca比等,可以重建鰻魚過去的洄游環境史。
GMM (Application of Genetic and Microchemical Maker as implements for diadromous and endangered commercial fish species populations management, 2004-2007) 計畫, 是行政院國家科學委員會支持的台灣-立陶宛-拉托維亞三國的國際合作計畫。自從1970-80年之後,全世界的溫帶鰻(日本鰻、美洲鰻及歐洲鰻)的資源量逐年下降。立陶宛和拉托維亞兩國地處歐洲鰻分布的邊疆地帶,境內天然洄游來的歐洲鰻明顯減少,於是從英國及法國進口鰻線來放流以便增加鰻魚產量。
但是放流之後,他們根本無法辨別何者是天然鰻,何者是放流的鰻,因而無法進行放流效益評估。我們利用中研院地科所的電子微探儀,測量耳石Sr/Ca比的時間變化,就能識別何者是天然鰻、何者是放流鰻。天然鰻因游過波羅的海,其耳石Sr/Ca比,從鰻線階段(EC)之後有一段高於千分之四的訊號(波羅的海訊號),而且冬季年輪形成時的耳石Sr/Ca比又比其他季節高,表示有季節性陸海移動現象(圖1.5.
79)。
放流鰻,因為是從英國或法國買來放流的鰻線,沒有游過波羅的海,其耳石Sr/Ca比,從鰻線階段(EC)之後都在千分之四以下,而沒有出現波羅的海訊號,因此很容易識別。換言之,耳石Sr/Ca比可以用來區別立陶宛和拉托維亞兩國境內的鰻魚,何者是天然鰻,何者是放流鰻(Shiao
et al. 2006. Lin
et al. 2012c)。
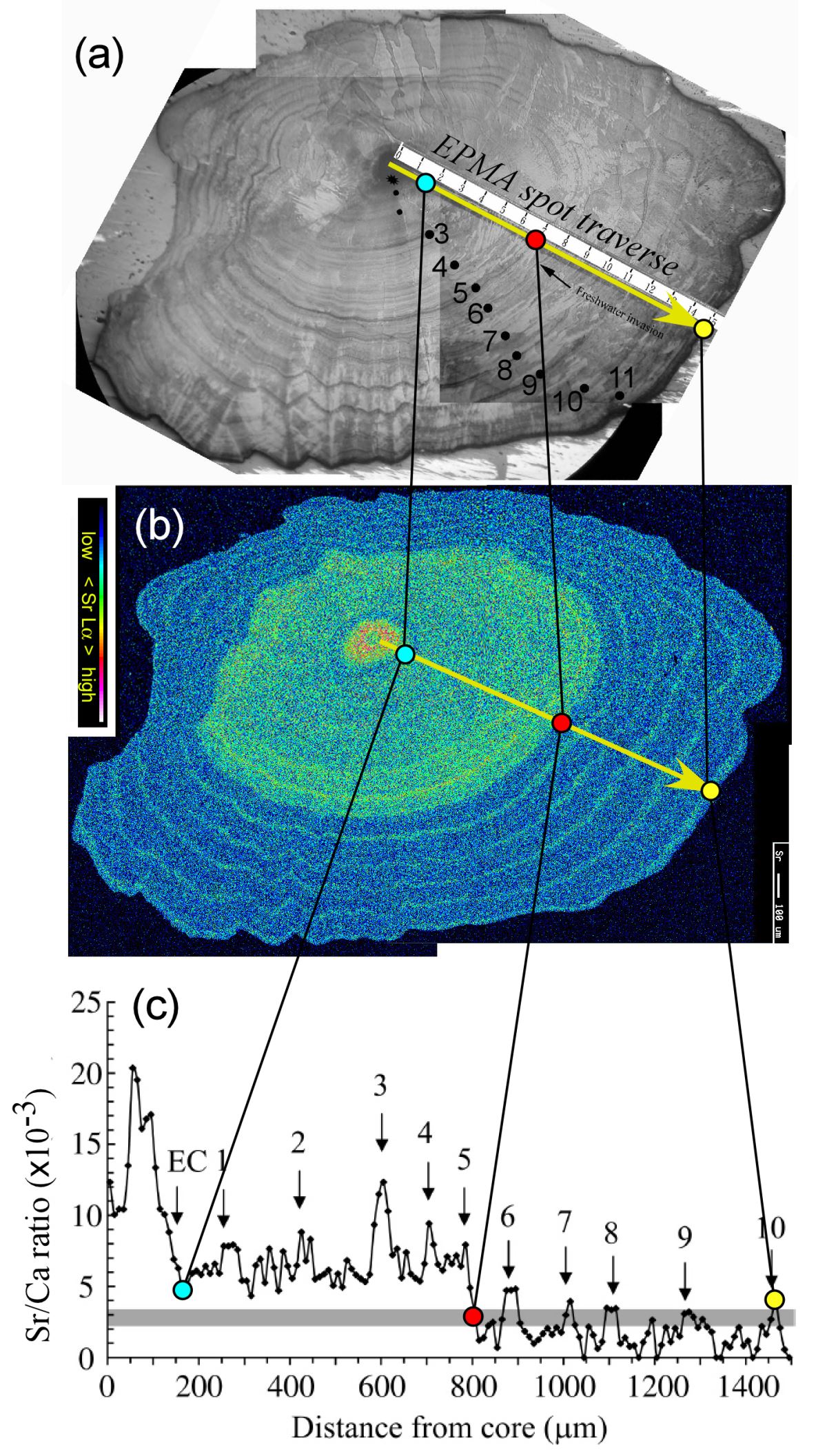
圖1.5.9天然歐洲鰻耳石的年輪(a.b)、Sr濃度分佈圖(b)以及Sr/Ca比從核心到耳石邊緣的變化(c, 數字代表年齡) (photo: Yoshiyuki Iizuka)
耳石碳酸鈣的結晶型有時,不知什麼原因,會從Aragonite變成Vaterite的同分異構物,陽離子的配位數也從9變為6時,有效離子半徑增加,兩價的Sr進入Vaterite的晶格受到限制,這時外在環境中即使有很高的Sr濃度,也不易進入耳石,因此利用耳石Sr/Ca比來識別其洄游環境時要留意耳石結晶構造(Tzeng
et al. 2007)。耳石就像是百寶箱一樣,蘊藏很多魚類生活史的祕密,隨著研究工具的發展,這些鮮為人知的祕密一一浮現。耳石的生態應用是跨領域的研究,必需結合生物地化的知識,魚類生態應用研究的路才會更寬廣。
1.5.7鰻魚洄游的進化(The evolution of eel migration)
耳石Sr/Ca比的證據顯示,日本鰻出現三種洄游環境史(圖1.5.
810): 1) 玻璃鰻到了沿岸之後不進入淡水河川,而在海水中生長,此稱之為海水型(Seawater contingent),其耳石Sr/Ca比在鰻線階段以後(即耳石半徑超過約150 µm以上)都維持在4 ‰ 以上,可予以證明,耳石Sr/Ca比4 ‰ 是淡水和海水的界限(Tzeng 1996); 2)淡水型 (Freshwater contingent),是鰻魚最典型的洄游模式,耳石Sr/Ca比的訊號在鰻線階段以後都維持在4 ‰ 以下; 3) 河口型鰻魚(Estuarine contingent)在淡水和海水環境之間移動,其耳石Sr/Ca比在4 ‰ 上下變動(圖1.5.
911) (Tzeng
et al. 2002)。換言之,成長期的黃鰻之棲地利用是逢機洄游性(Facultative catadromy or Semi-catadromy)而非強迫洄游性(Obligatory catadromy)(Tsukamoto and Arai 2001,Tzeng
et al. 2001)。這樣的洄游進化,可以降低河川棲地喪失、或河川污染時所帶來的風險,例如某河川因淡水截流或污染而無法溯河時,鰻魚仍然可在海水域成長。海水型鰻魚的比例,有隨緯度的增加而增加的趨勢,這可能是高緯度的海水生產力比淡水高的原因(Deverat
et al. 2006)。由此可見鰻魚棲地利用的可塑性很大。
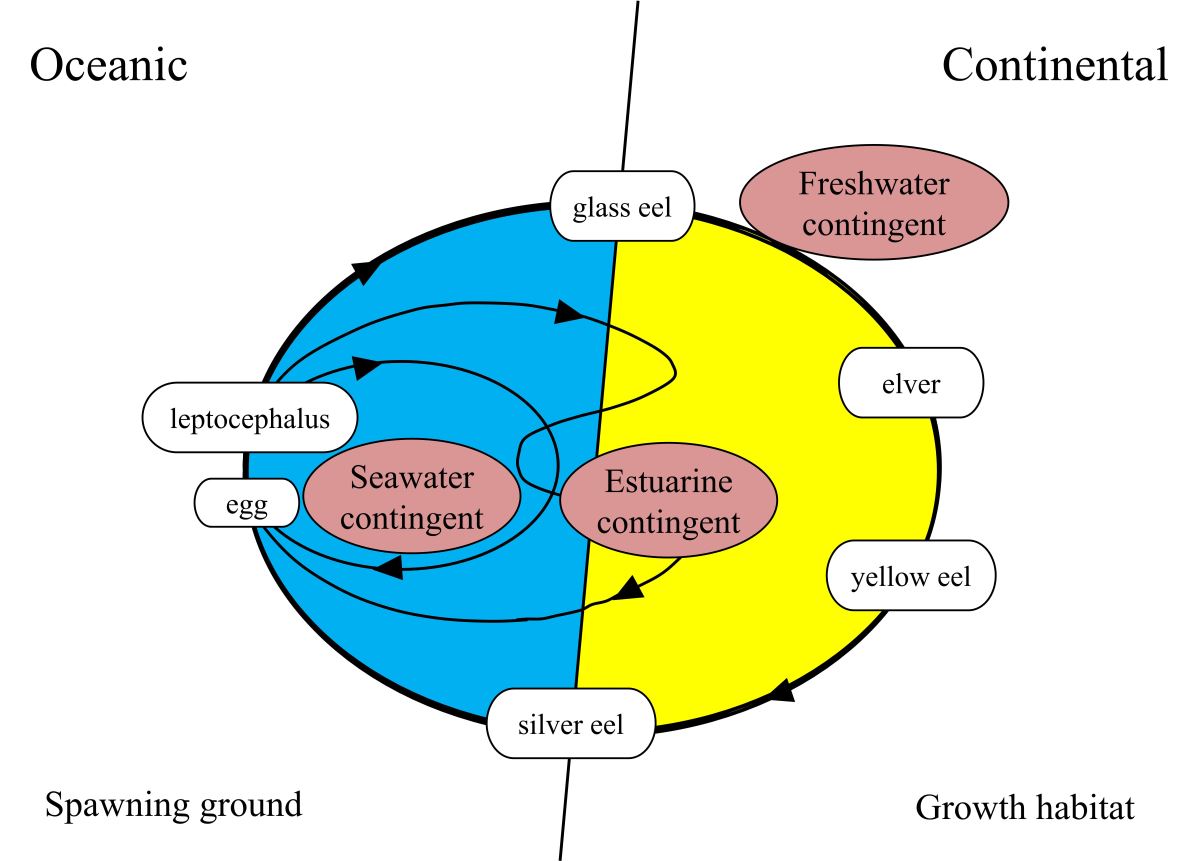
圖1.5.
10三種生活史的洄游模式示意圖:1) 海水型(Seawater contingent),2)淡水型(Freshwater contingent),3)河口型(Estuarine contingent)
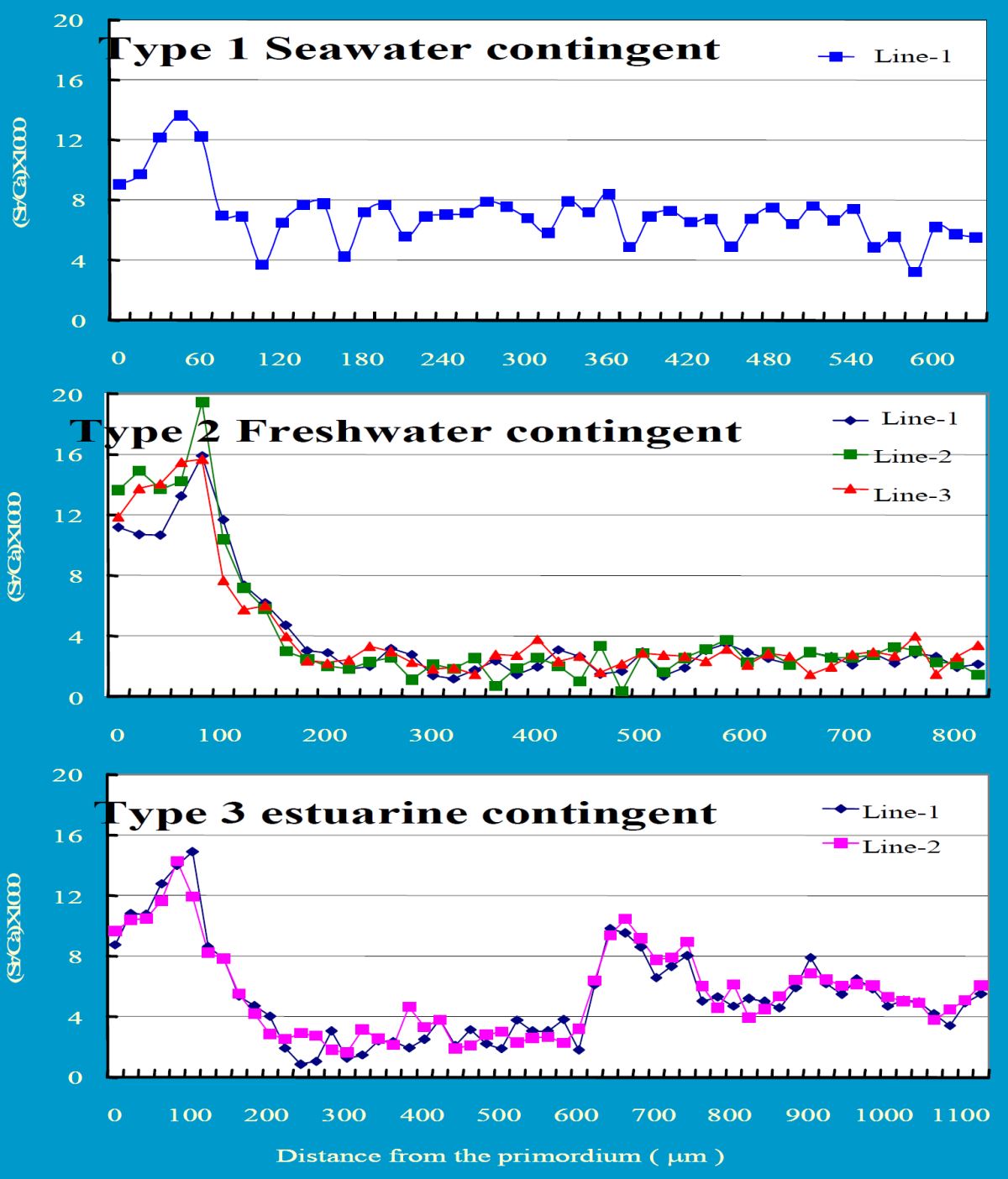
圖1.5.
11魚三種洄游模式的耳石Sr/Ca比之比較。Type 1: 海水型(Seawater contingent), Type 2: 淡水型(Freshwater contingent),Type 3: 河口型(Estuarine contingent)
1.5.11鰻魚耳石研究的合作國家(Countries collaborating with Taiwan for the eel otolith study)
臺灣與世界各國合作研究鰻魚耳石的國家有亞洲的中國大陸、日本、菲律賓;美洲的加拿大、美國、海地;歐洲有立陶宛、拉托維亞、瑞典、英國、愛爾蘭、法國、葡萄牙、義大利及土耳其等;非洲有南非和馬達加斯加;大洋洲有澳洲和紐西蘭等十九個國家(圖1.5.
12)。合作研究的鰻魚種類有日本鰻、鱸鰻、美洲鰻、歐洲鰻、莫三比克鰻、澳洲鰻以及紐西蘭大鰻等,發表的學術論文多達 30篇 (例如Briones
et al. 2007; Cairns
et al. 2004; Daverat et al. 2006; Lamon
et al. 2006; Lin
et al. 2007, 2009, 2010, 2011, 2012a,b,c; Jessop
et al. 2002, 2006, 2007, 2008a,b, 2011; Panfili
et al. 2012; Shiao
et al. 2001a,b, 2002, 2006; Tabeta
et al. 1987; Thibault et al. 2007; Tzeng
et al. 1994, 1997, 1999, 2000, 2003, 2005, 2007)。合作研究項目有耳石日周輪和耳石微化學(包括耳石Sr/Ca比和微量元素組成)。從耳石日周輪研究瞭解到鰻魚的出生日、變態日齡及到達河口的日齡等初期生活的演化策略,從耳石微化學研究瞭解到鰻魚的洄游環境及棲地利用的多樣性。這些國際合作,要特別感謝行政院國家科學委員會的長期補助研究經費、國際合作伙伴以及臺灣大學生科院漁業科學研究所和理學院動物系的碩、博士生的奉獻,才能建立台灣鰻魚耳石研究的國際學術知名度。曾教授的耳石研究團隊從1990年到2012年總共發表一百多篇鰻魚論文(詳臺灣大學圖書總館臺大人文庫的曾教授退休論文集及鰻魚生活史及保育論文集)。
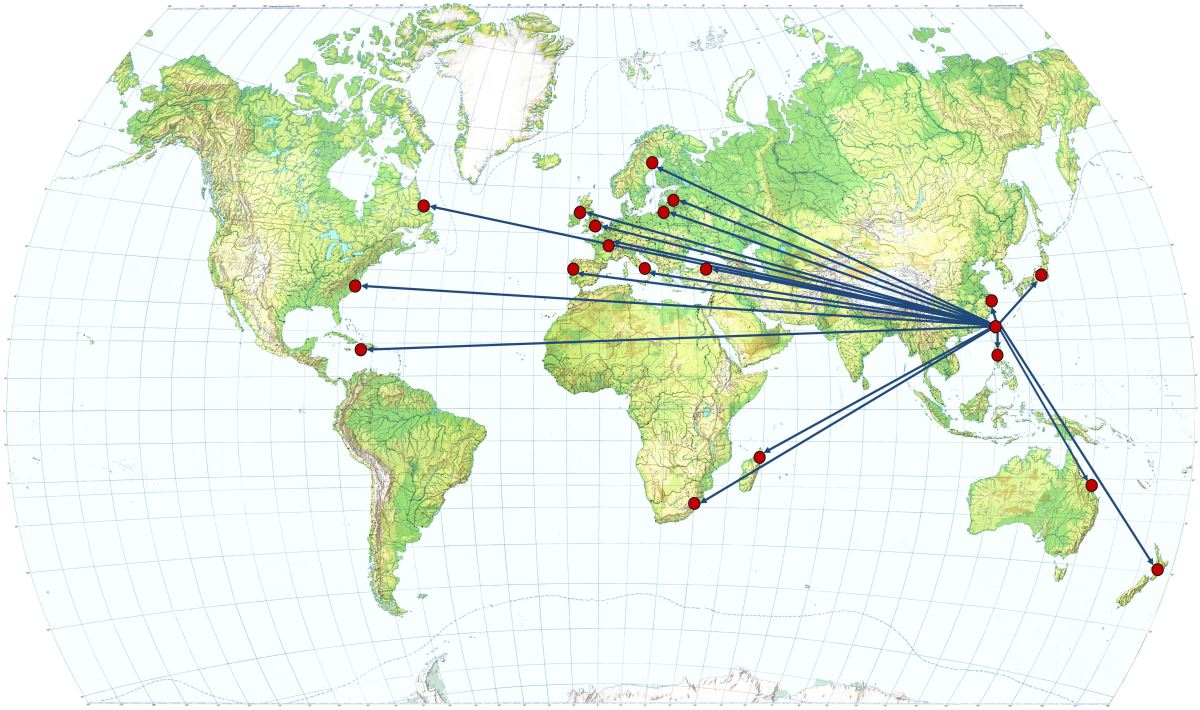
圖1.5.
12與臺灣合作研究鰻魚耳石的國家