洄游是魚類最基本的生存策略。洄游比不洄游更容易獲得資源而繁衍更多子代。本章將探討兩側洄游性魚類的進化、日本鰻洄游史、以及鰻魚誕生之後如何漂洋過海上溯到陸地河川的成長棲地。並且介紹追蹤銀鰻降海產卵洄游的最新科技。
1.4.1 兩側洄游性魚類的進化(The evolution of diadromons fishes)
依棲息水域不同,魚類分為淡水魚類(Freshwater fishes)、海水魚類(Marine fishes)、以及在陸地河川和海洋之間洄游的兩側洄游性魚類(Diadromons fishes)(圖1.4.1)。兩側洄游性魚類又分為:(1)在海洋中生長而上溯到河川上游產卵的溯河洄游性魚類(Anadromons fishes),例如鮭魚;(2)在河川中生長而降河洄游到海洋產卵的降海洄游性魚類(Catadromons fishes),例如鰻魚(
Anguilla spp.); 以及(3)不是為產卵而在陸地河川和海洋之間洄游的兩棲洄游性魚類(Amphidromous fishes),如蝦虎魚(Goby)。
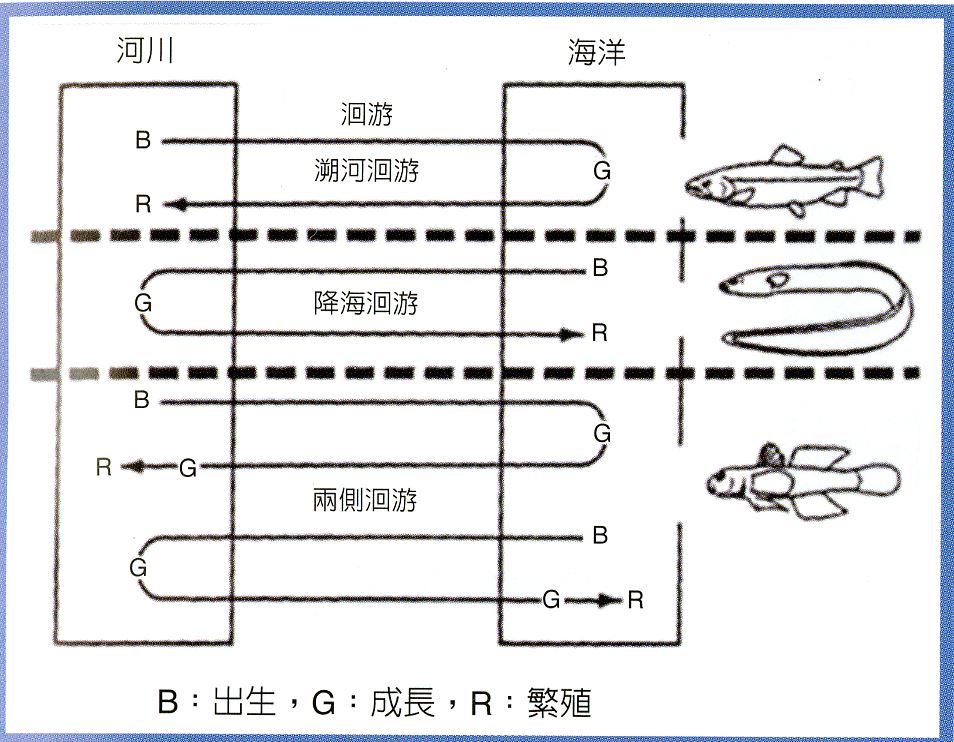
圖1.4.1河海之間洄游的魚類有: 溯河產卵洄游型(Anadromy)的鮭魚、降海產卵洄游型(Catadromy)的鰻魚以及兩棲洄游型(Amphidrony)的蝦虎魚。
根據Gross(1988)的研究發現,高緯度地區的水域生產力,河川低於海洋,而低緯度地區則海洋低於熱帶雨林的河川。魚類為了快速成長及繁衍更多子代,選擇洄游到餌料生物豐富的地方生長,而洄游到敵害相對較少的貧營養水域產卵,以便降低卵及仔魚被捕食的風險。於是演化出高緯度地區的鮭魚,選擇在低生產力的河川產卵而洄游到高生產力的海洋生長。反之,低緯度地區的鰻魚則選擇低生產力的熱帶海域產卵而洄游到高生產力的熱帶及溫帶地區的河川生長。兩者的演化有異曲同功之妙。
1.4.2 日本鰻的洄游史(The migratory history of the Japanese eel)
鰻魚的生活史分為七個發育階段:卵(Egg stage)、前期柳葉鰻(Pre-leptocephalus)、柳葉鰻(Leptocephalus)、玻璃鰻(Glass eel)、鰻線(Elver)、黃鰻(Yellow eel)以及銀鰻(Silver eel)(圖1.4.2)。日本鰻在陸地河川中生長5-6年後,便從黃鰻階段變態為銀鰻,洄游到海洋中產卵,體色的改變是為了適應降河洄游產卵時的深海環境。一生只產一次卵。產完卵後就葬身海底結束其傳奇的一生。日本鰻產卵場與陸地相隔5000多公里,花半年時間長途跋涉到大洋區的深海產卵場產卵,這是生物演化的一項奇蹟。大洋區生產貧瘠,天敵少,仔代活存率高,再藉著海流分散風險將子代分散到東北亞國家的台灣、中國大陸、韓國及日本等地的內陸河川的高生產區生長。這種巧妙的生活史策略,使得鰻魚在地球上不容易滅絕的原因。
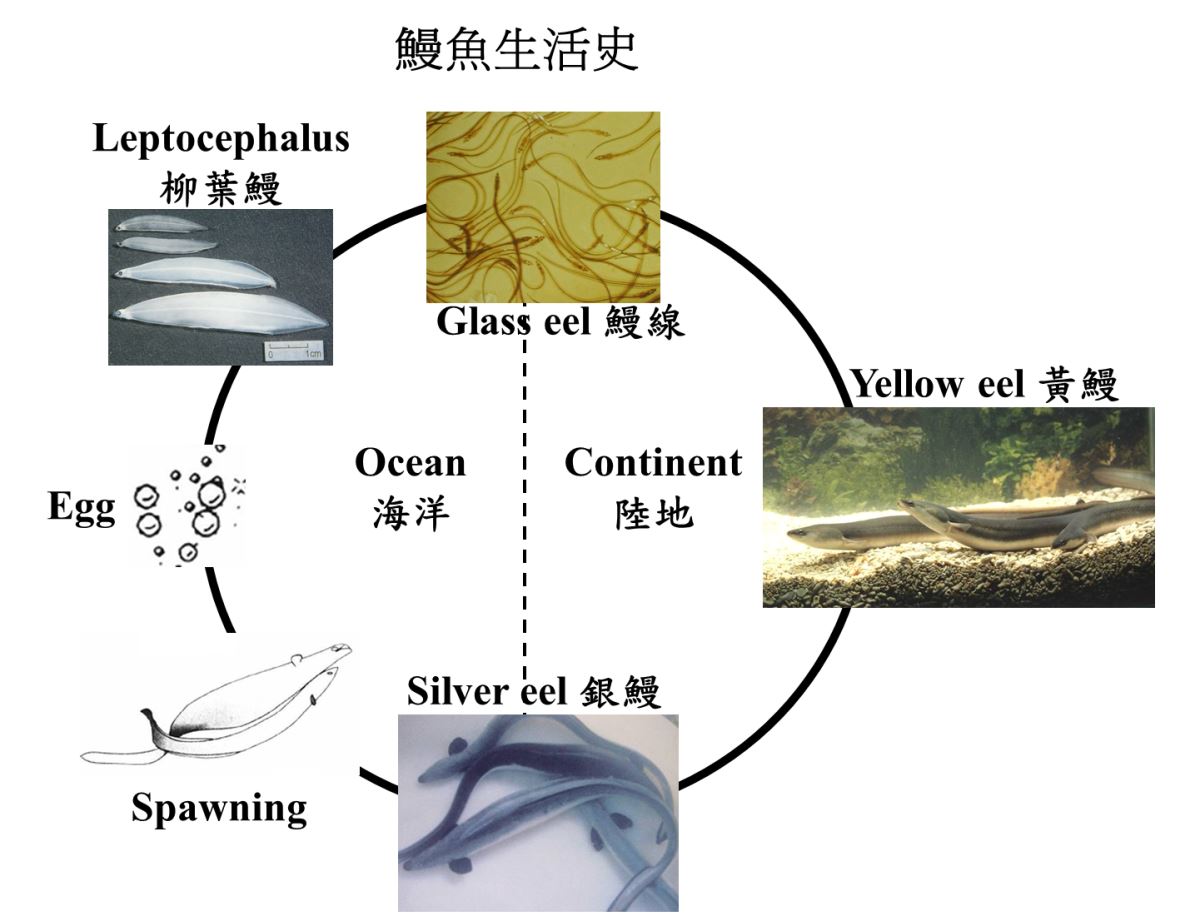
圖1.4.2鰻魚生活史及其發育階段(卵、柳葉鰻、玻璃鰻、黃鰻以及銀鰻)
- 柳葉鰻的洄游路徑(The pathway of larval migration from ocean to estuary )
近三、四十來日
本研究團隊從海上採集的柳葉鰻體型大小的時空分布,可約略拼湊出日本鰻的產卵場位置以及柳葉鰻的洄游路徑(圖1.4.3)。日本鰻每年4~11月在馬里亞納群島西方海域產卵。孵化之後的前期柳葉鰻和柳葉鰻,體型側扁、狀如柳葉,適合海上的長距離漂游,隨北赤道洋流由東往西漂,到了菲律賓東方海域後北轉進入黑潮。11~3月接近東北亞國家大陸棚時柳葉鰻變態成為流線型的玻璃鰻,脫離黑潮進入沿岸水域。
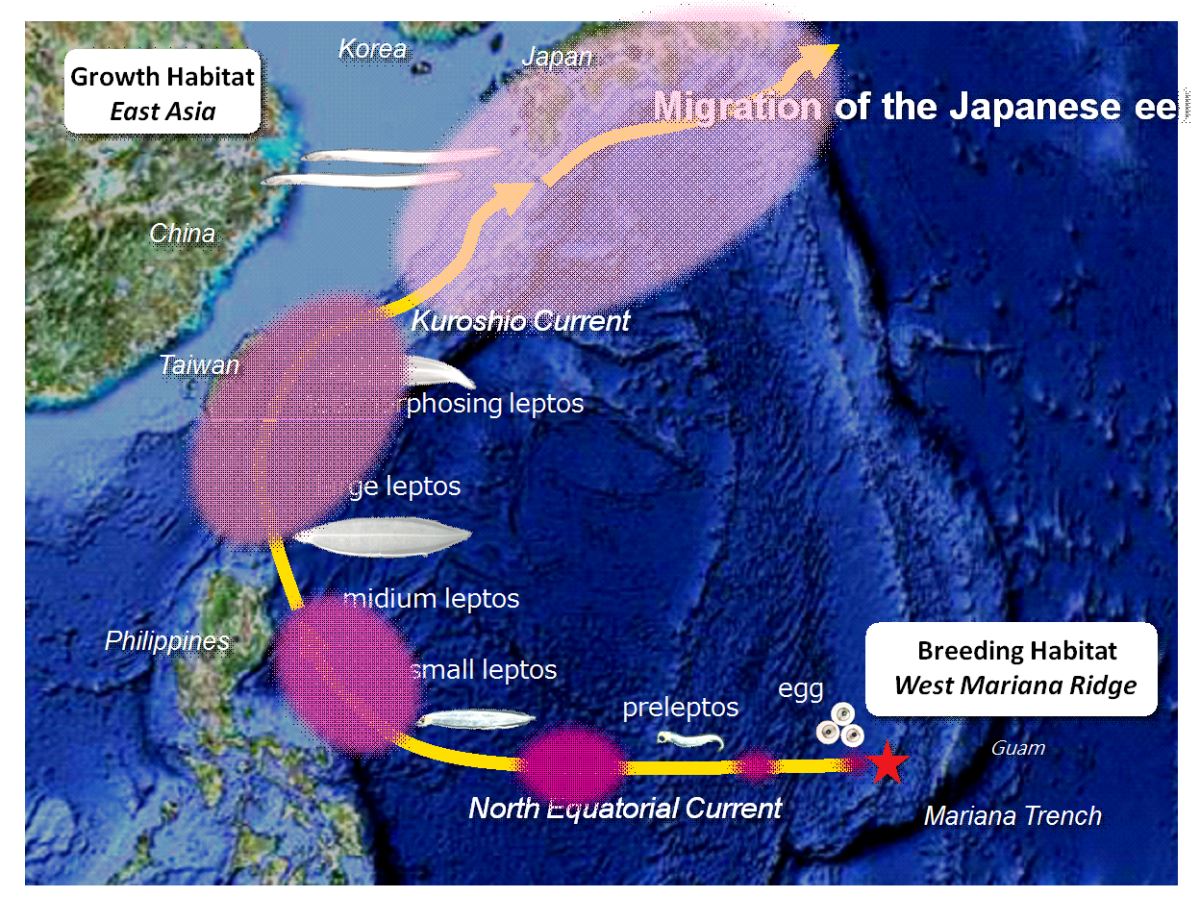
圖1.4.3日本鰻在馬里亞納海溝西方海域產卵,孵化之後的柳葉鰻順著北赤道洋流與黑潮漂送到東北亞國家大陸棚時變態成為流線型的玻璃鰻,然後脫離黑潮進入沿岸水域。(photo: Katsumi Tsukamoto)
(2) 海流對柳葉鰻漂送路徑的影響(The effect of current on larval dispersal )
日本鰻誕生之後的向岸洄游,可分兩階段。第一階段是柳葉鰻順著北赤道洋流由東向西漂送,到了菲律賓外海之後,北轉進入黑潮經過台灣、中國大陸、韓國及日本等國家外海。第二階段是柳葉鰻變態成玻璃鰻之後,脫離黑潮進入上述國家的沿岸水域。在北赤道流的輸送過程中,進行日周垂直移動,白天在200公尺左右,晚上則上昇到100公尺左右(Otake
et al. 1998)。這種特殊的行為,一來可以減低被掠食者(例如燈籠魚)捕食,二來是為了確實追隨餌料生物的移動。
柳葉鰻夜間浮到表層也為了從北赤道洋流北轉進入黑潮(Kimura
et al. 1994)。如果沒有進入黑潮很可能南下進入民答那娥海流而被輸送到熱帶海域,造成輸送到東北亞地區的玻璃鰻變少(Kim
et al. 2007)。產卵場的位置若南移,會造成柳葉鰻從產卵場輸送到東北亞國家的數量變少。日本鰻在14
o N 142
o E海域產卵,大部分的柳葉鰻都能從北赤道洋流進入黑潮,而來到東北亞國家。若產卵場往南偏一度(13
o N 142
o E)的話,大部分的柳葉魚會從北赤道洋流進入民答那娥海流而輸送到本來沒有日本鰻分布的印尼海域(Kuroki
et al. 2009)。產卵場的位置僅僅1度之差,柳葉鰻最終的漂送地點,就可能有天壤之別(圖1.4.4)。
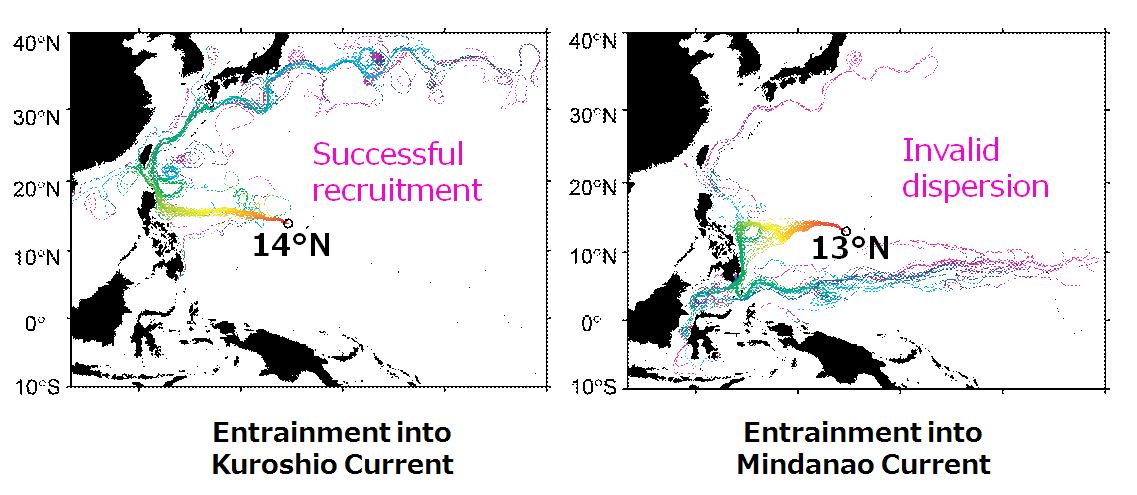
圖1.4. 4產卵場位置改變時柳葉鰻漂柳葉鰻的洄游路徑過程的數值模擬結果(Photo: Mari Kuroki,Heeyong Kim and Shingo Kimura)
另一方面,Han
et al.(2012)認為台灣地區的日本鰻玻璃鰻,大部分隨著黑潮主流經由台灣東部由南向北進入宜蘭和北北基東部地區,小部分則藉由黑潮支流(TSC)經巴士海峽進入台灣海峽沿著台灣西海岸北上(圖1.4.5)。
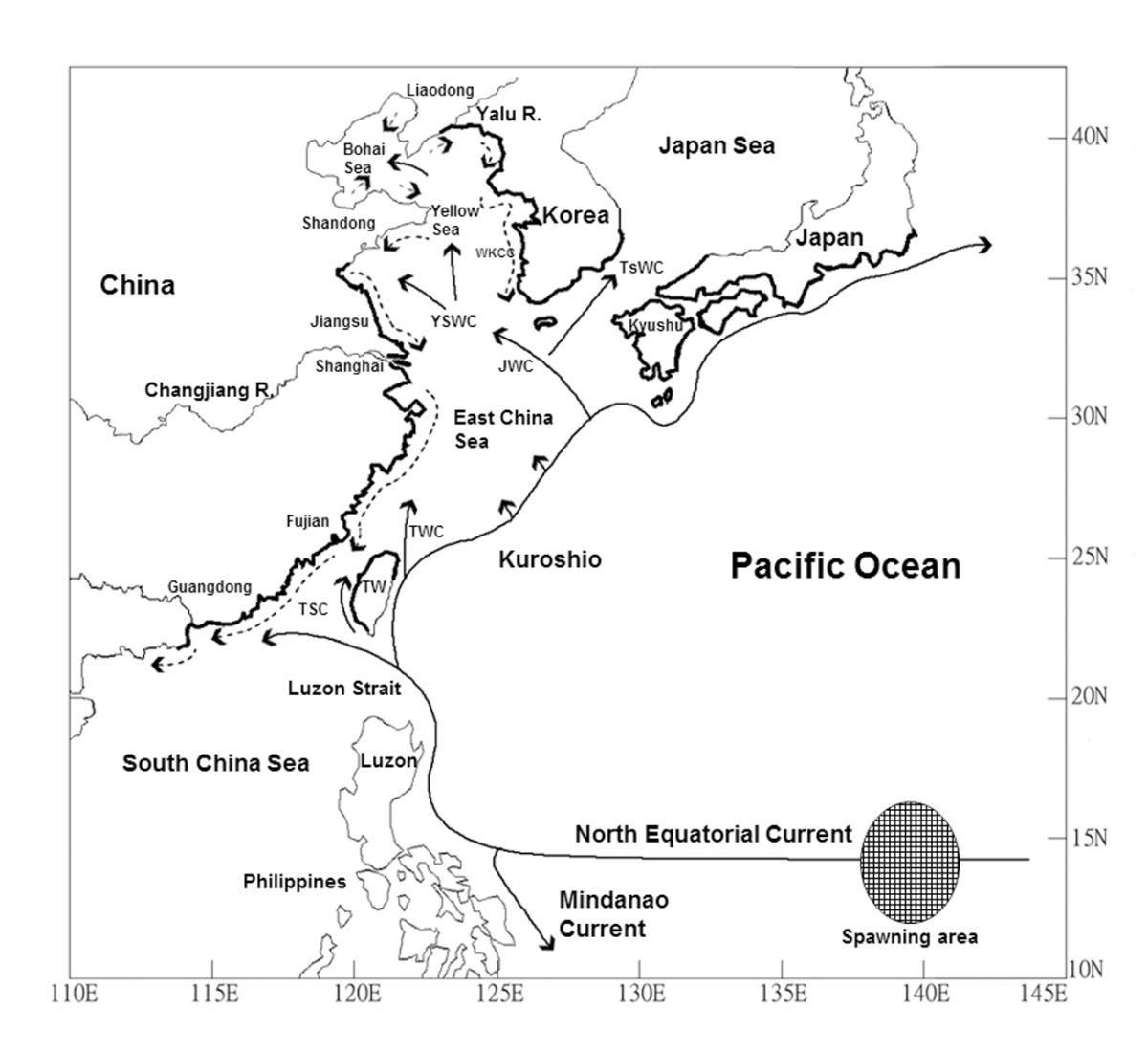
圖1.4.5日本鰻玻璃鰻到了西北太平洋國家的大陸棚之後隨著黑潮 及其支流(TSC,TWC,JWC,YSWC,TSWC)進入沿岸(黑色粗線部分)。
1.4.3 玻璃鰻的溯河行為和環境誘因(The upstream migratory behavior of glass eel and environmental cues for migration)
台灣沿岸的日本鰻玻璃鰻是東北季風增強之後隨中國大陸沿岸流南下到台灣東北部和西部的。以台灣東北部的福隆地區為例:中國大陸沿岸流南下最強,水溫降低到冬季平均最低水溫(約15
oC)時,玻璃鰻就順著漲潮流大量湧入河口域,於是玻璃鰻的漁獲量和單位努力漁獲量今均顯著增加(圖1.4.6。玻璃鰻剛剛到沿岸時,皮下黑色素胞還沒有發育完全很怕光。因此,農曆十五大潮有月光時,大都潛在沙底不活動(圖1.4.7,不易捕捉而漁獲量就變少(Tzeng 1985)。
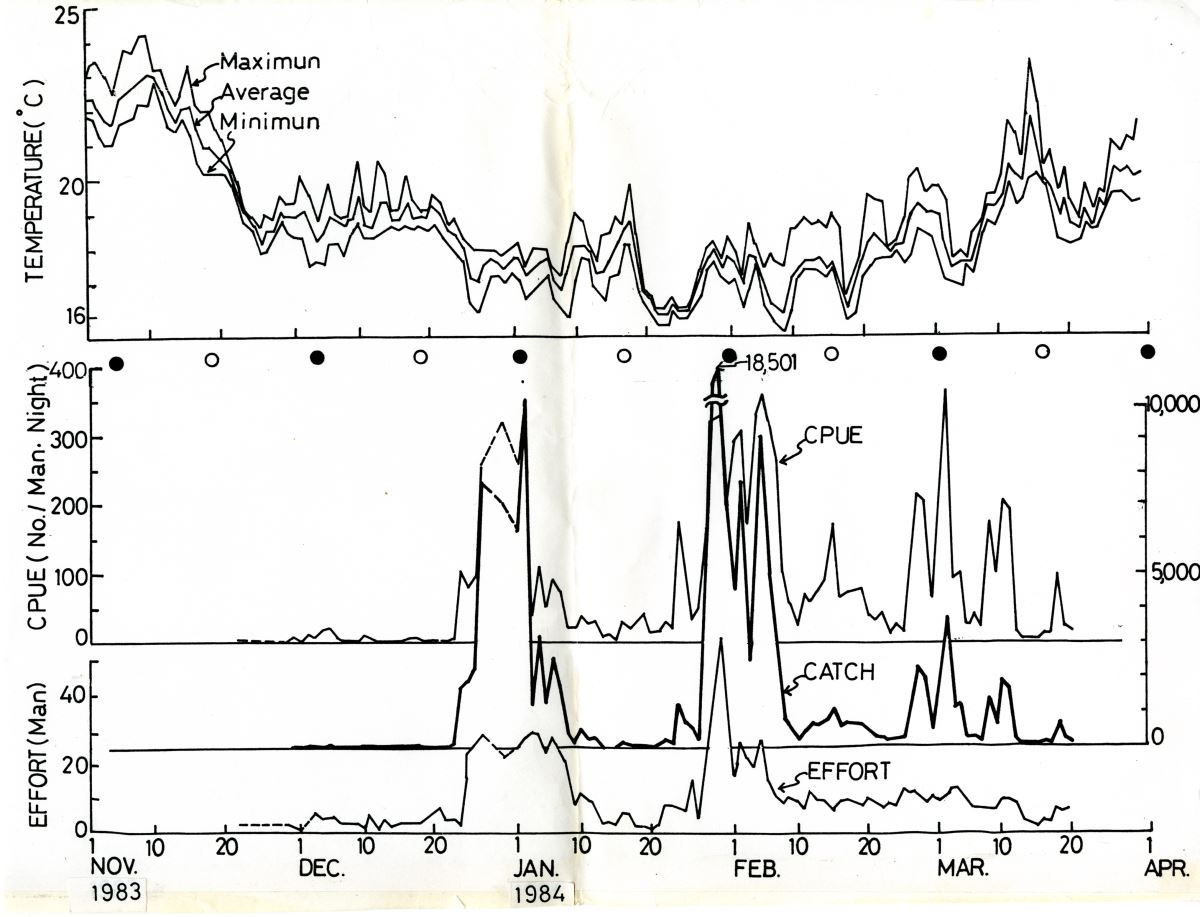
圖1.4.6鰻線的漁獲量(Catch)、努力量(Effort)和平均漁獲量(CPUE, 每人每晚的捕獲尾數)的日變化與水溫、月亮週期之關係。
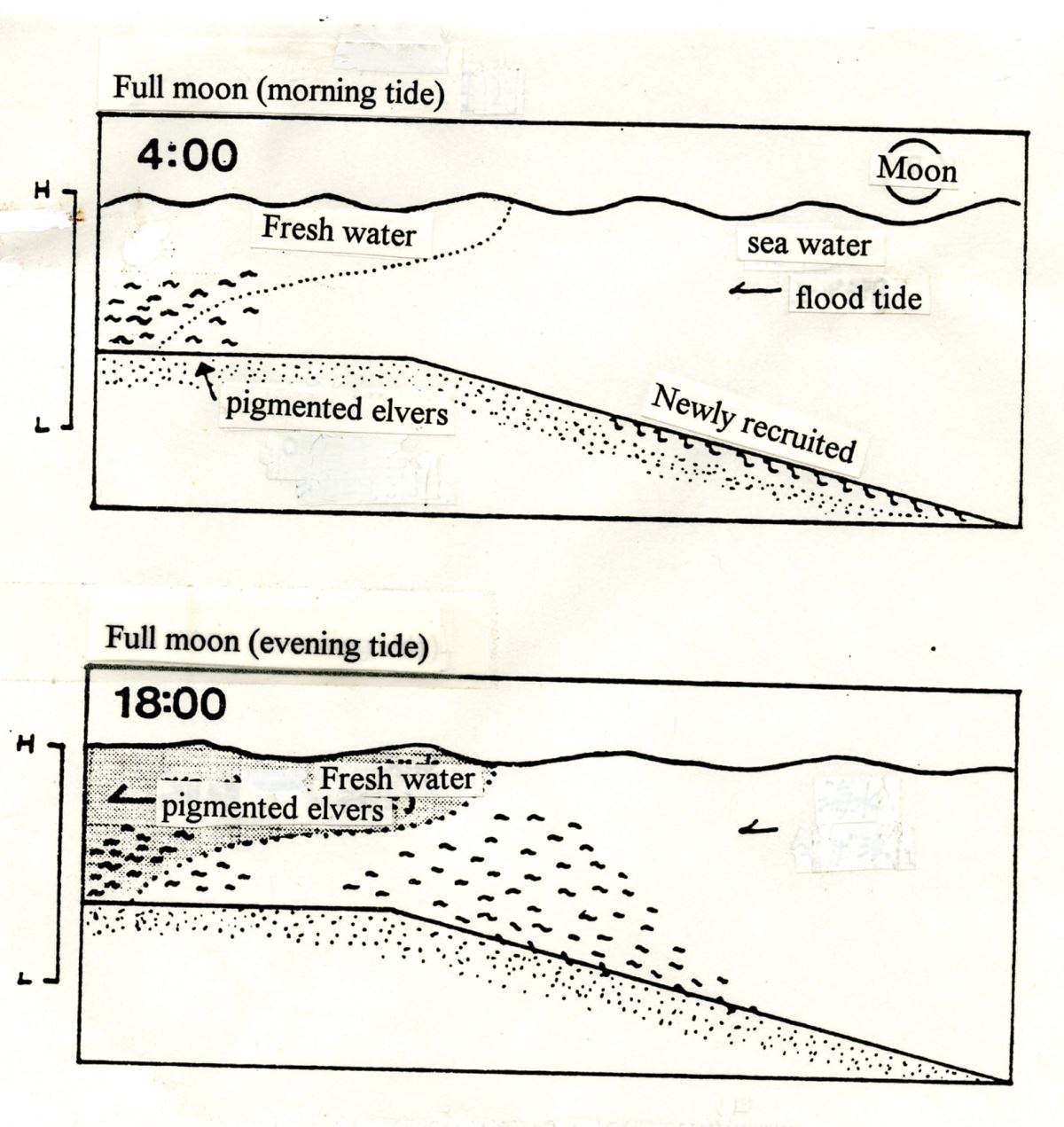
圖1.4.7玻璃鰻的溯河行為。(上圖)滿月時,剛剛到達河口的玻璃鰻(Newly recruited glass eels)發育階段為VA,頭頂尚未長出黑色素胞,對光線敏感而埋藏在沙泥中,即使漲潮也不溯河; 但發育階段為VIA,已經長出黑色素胞的鰻線(pigmented elvers)則不受月光影響,隨漲潮溯河。 (下圖)無月光時,玻璃鰻也順著漲潮流溯河(modified from Sakurai 1975)。
但進入河川後,玻璃鰻黑色素胞逐漸發育到VIA階段的鰻線,鰻線對光就沒有那麼敏感,河內在有月光出現的農曆十五大潮時,鰻線也會順著漲潮流溯河,因此農曆初一和十五的大潮皆有很高的漁獲量(圖1.4.8)(Tzeng 1985)。
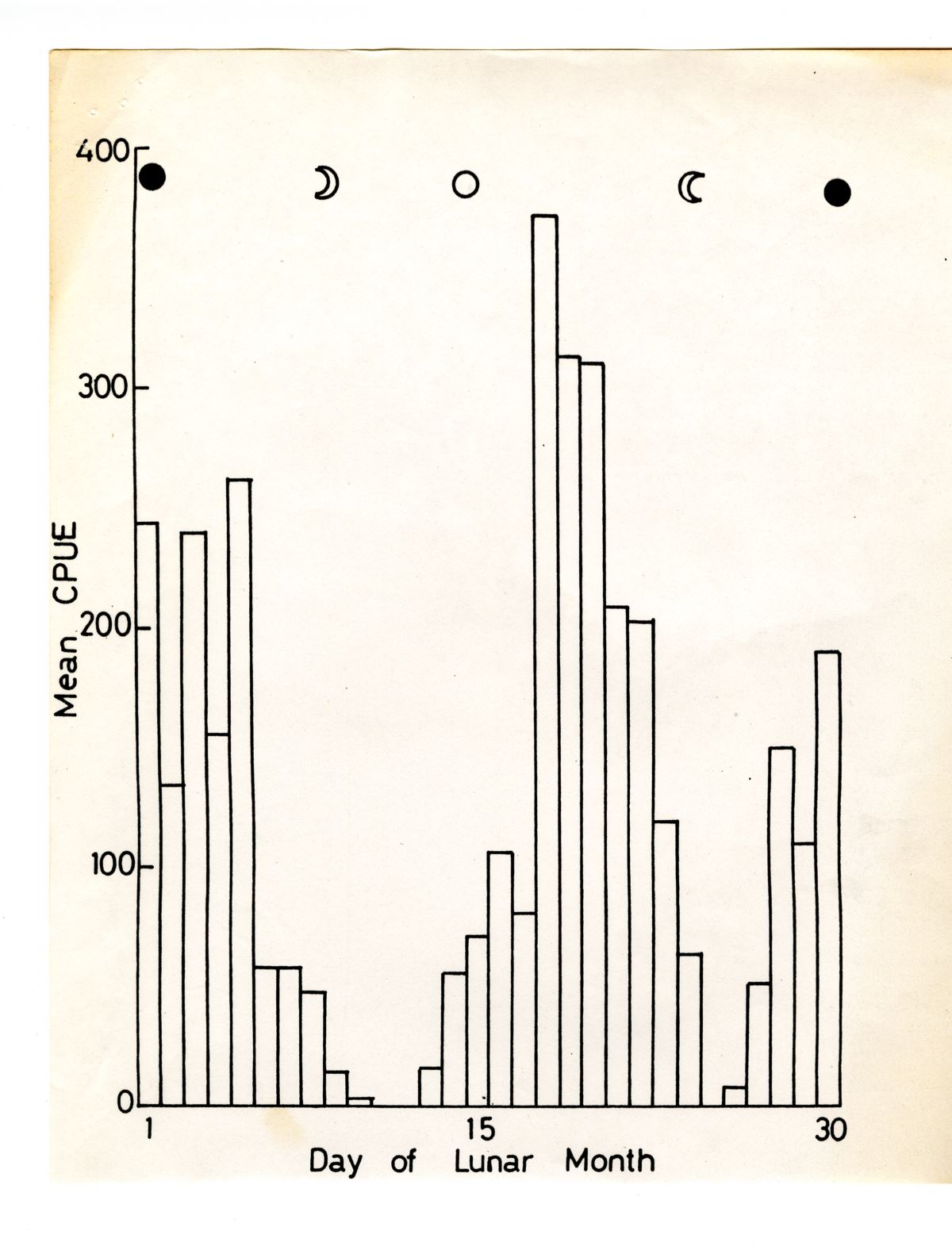
圖1.4.8東港溪1981-1982年冬季 鰻線的平均漁獲量(CPUE, 每人每晚的捕獲尾) 的日變化與月週期之關係。
1.4.4 銀鰻降海產卵洄游的追蹤技術(Techniques for tracking the seaward spawning migration of silver eels)
黃鰻在河川成長4-6年後,生殖腺逐漸發育而變態成為銀鰻。銀鰻腹側為銀灰色,降海前消化道開始退化不再進食,同時眼睛變大,胸鰭變黑變寬,以便適應深海產卵洄游。每年秋冬大雨過後銀鰻會進行降海洄游產卵。銀鰻從陸地河川的成長棲息地回到產卵海域的洄游路徑尚不清楚。科學家曾經利用浮上式衛星標簽(PSATs, pop-up satellites achival tags),追蹤愛爾蘭的歐洲鰻(Aarestrup
et al. 2009)或紐西蘭大鰻的降海洄游行為(圖1.4.9)。但都因追蹤的日數非常短,無法窺知銀鰻從陸地河川回到產卵場的全程洄游路徑(Jellyman and Tsukamoto 2010)。

圖1.4.9a Don Jellyman 博士正在用尼龍線把衛星標籤固定在一條1.3公尺麻醉中的雌性紐西蘭大鰻(
A. dieffenbachii)身上(photo: Don Jellyman)

圖1.4.9b標上衛星標籤的大鰻,利用側面的白色圓盤固定尼龍線綁住背上的衛星標籤,當深度超過2000公尺時,衛星標籤與尼龍繩之間有自動切離裝置,以免衛星標籤受壓損壞(photo: Don Jellyman)

圖1.4.9c Don Jellyman 博士在紐西蘭的Canterburg省靠近Ellesmere湖的海邊釋放標識後的鰻魚(photo: Don Jellyman)
日本的研究團隊近年也在日本近海進行日本鰻的衛星標示追蹤器,追蹤銀鰻如何從陸地河川回到馬里亞納島西側的產卵場(圖1.4.10a、b、c)。初步發現銀鰻降海後,會在水溫5
o-15
oC,深度200-600 m之間進行規律性的日夜深淺洄游(Manabe et al. 2011)(圖1.4.10 d)。可是,根據美國Woods Hole海洋研究所愛爾文號(Alvin)潛航器在藻海附近的Bahamas 島海域的觀測,非常驚訝地發現美洲鰻銀鰻的洄游深度可能深達2000m(Robins
et al. 1979)。至於日本鰻銀鰻是否順著黑潮或其他路線回到馬里亞納島西側的產卵場,因衛星標示追蹤器的電池壽命有限,追蹤日數太短,其降海產卵的洄游路線仍然是個謎(圖1.4.10e)(Tsukamoto
et al.未發表)。
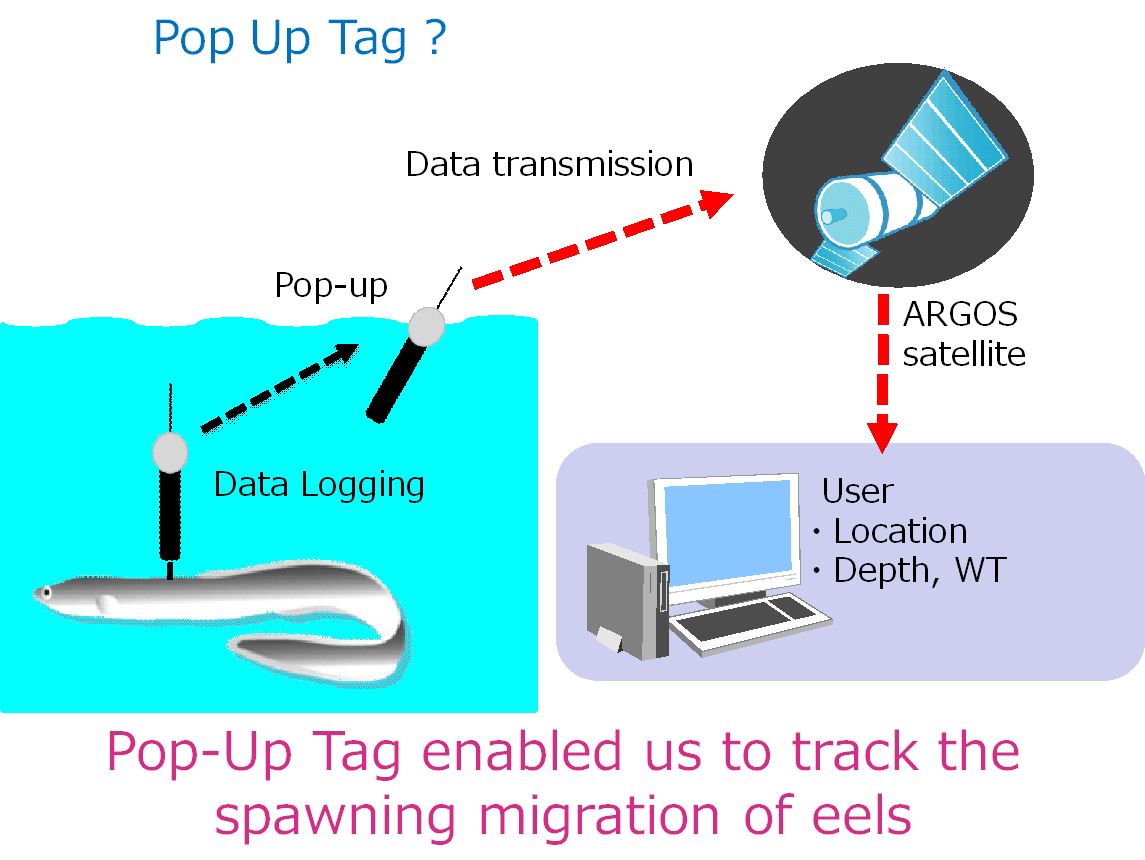
圖1.4.10a浮上式衛星標籤(pop-up satellite archival tag)到了設定時間就脫離魚體浮到海面將標籤說記錄的資訊透過衛星傳回電腦

圖1.4.10b銀鰻的浮上式衛星籤的標示過程(photo: Katsumi Tsukamoto)
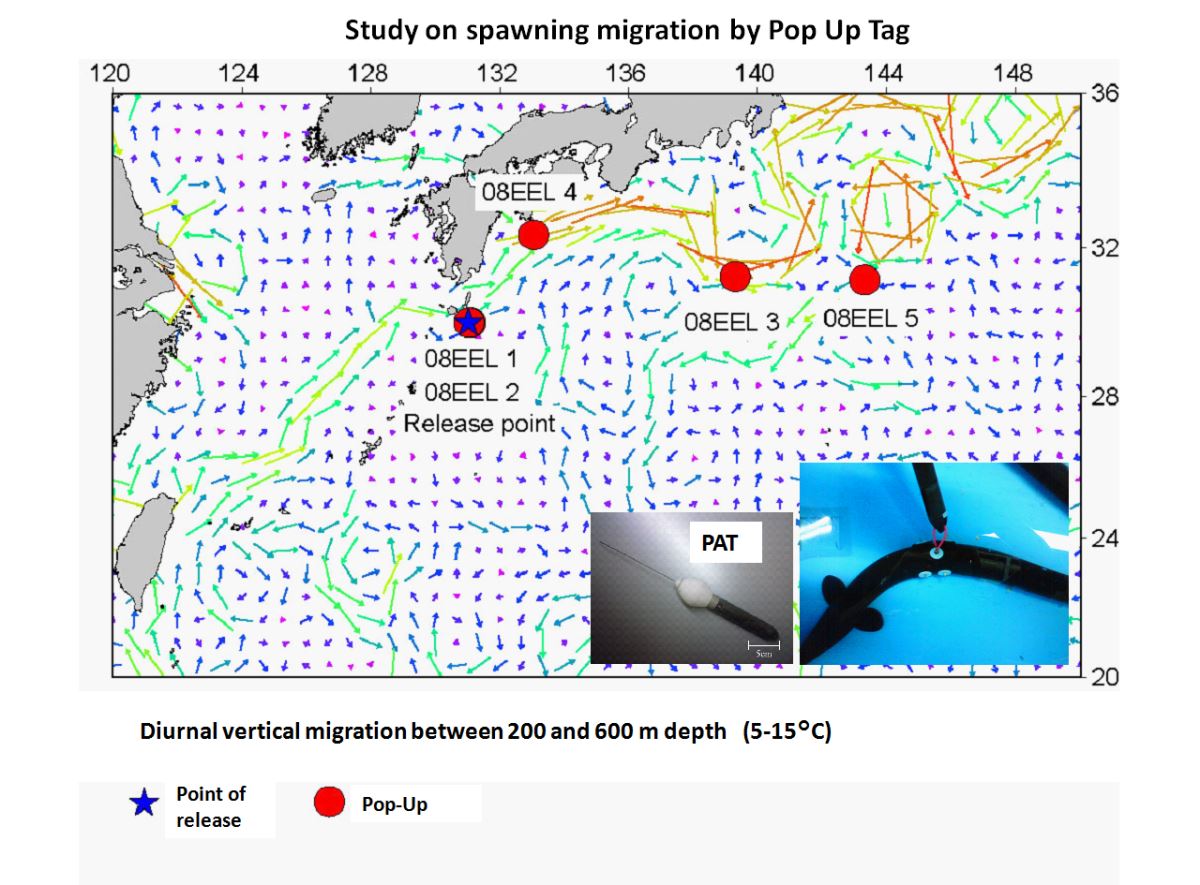
圖1.4.10c利用浮上式衛星標示法追蹤日本近海日本鰻銀鰻的降海洄游行為(Manabe
et al. 2011)
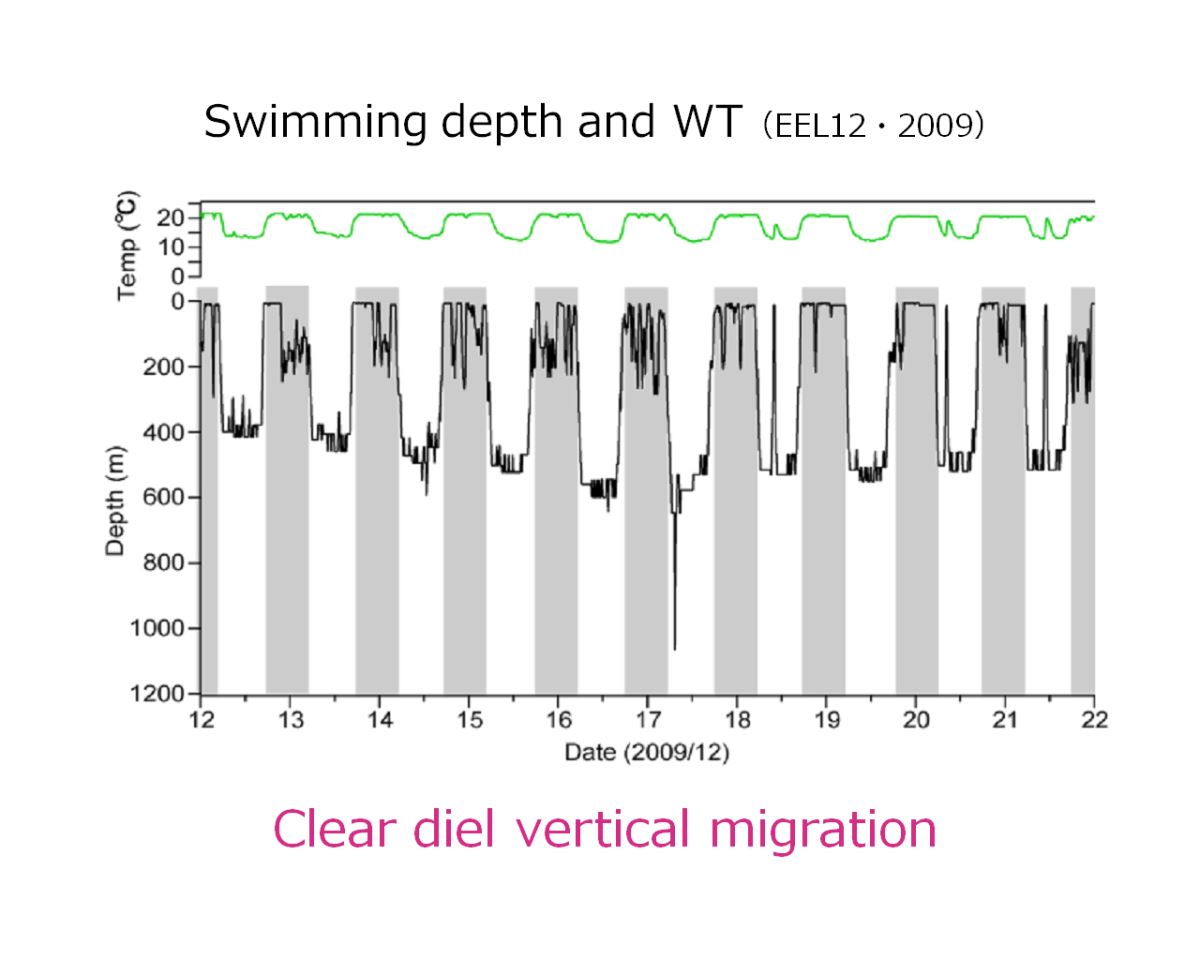
圖1.4.10d銀鰻降海的日夜深淺洄游行為(Manabe
et al. 2011)
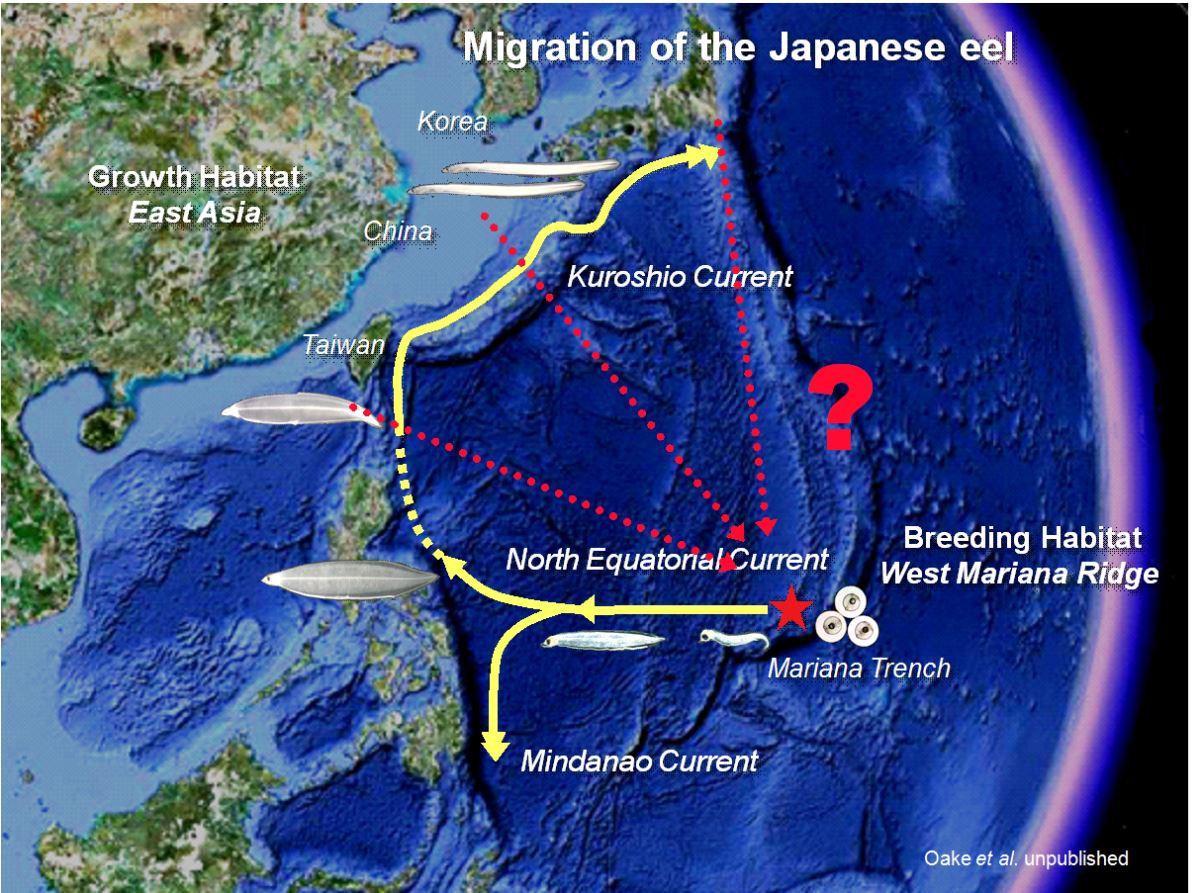
圖1.4.10e 銀鰻降海產卵的洄游路線仍然是個謎(photo: Katsumi Tsukamoto)
此外,為了觀察銀鰻的產卵場行為。1998年6月10~28日本研究團隊與德國的馬克斯普蘭克研究所(Maxplank Institute)合作,利用JAGO小型潛水艇潛航在馬里亞納島附近的日本鰻產卵場,沿著海山的斜坡,水溫10℃以下,深度400 m,上下來回27趟91小時,想一窺銀鰻聚集和產卵行為,但並未發現鰻魚蹤跡(圖1.4.11 a, b)。

圖1.4.11a日本東京大學海洋研究所的研究船”白鳳丸”與德國馬克斯普蘭克研究所(Maxplank Institute)的JAGO小型潛水艇(photo: Hans Fricke & Katsumi Tsukamoto)

圖1.4.11b JAGO小型潛水艇潛航到400 m 深(photo: Hans Fricke)