鰻魚的生命過程有很多不可思議的變化。每一階段的變化都有其環境適應和演化的道理。本章將從其生命誕生開始、柳葉鰻、溯河過程的玻璃鰻、河川內成長黃鰻及其性別決定、乃至降河產卵的銀鰻等階段的變化來瞭解鰻魚發育和成長的特徵。
1.3.1 生命從卵開始到前期柳葉鰻(Life starting from egg to pre-leptocephalus)
日本鰻在馬里亞納島西側海域產卵。卵的直徑約1.6mm,屬於分離式半浮性卵。卵受精2小時後可看到卵黃和油球各一(圖1.3.1)。受精卵隨波逐流漸漸分散。2009年5月22日(新月之前2天)天亮之前,日本的研究團隊曾經在馬里亞納海嶺南端的西側海山採到31個日本鰻的受精卵(圖1.3.2)。利用DNA鑑定結果確定為日本鰻無誤,因而驗證了日本鰻在海山附近產卵的假說。這是人類有史以來第一次採到野生的日本鰻受精卵。日本鰻是每年5、6月新月的晚上,於高鹽度和低鹽度水塊相接的海洋鹽度鋒與西馬里亞納海嶺的交匯處產卵的。
受精卵孵化前,約略可以看到眼睛和內耳的出現,心臟已開始跳動(圖1.3.3)。在25℃水溫下30小時就孵化。剛孵化時會激烈運動,完全沒有逃避敵害的能力,口未開,眼睛尚未出現色素,強光照射會導致失明(圖1.3.4)。

圖1.3.1人工養殖鰻的受精卵,直径約1.6mm(photo: Irago Institute)

圖1.3.2人類第一次在野外採到的天然日本鰻受精卵(Tsukamoto
et al. 2011)

圖1.3.3授精卵孵化前的胚胎發育情形(Tsukamoto
et al. 2011)
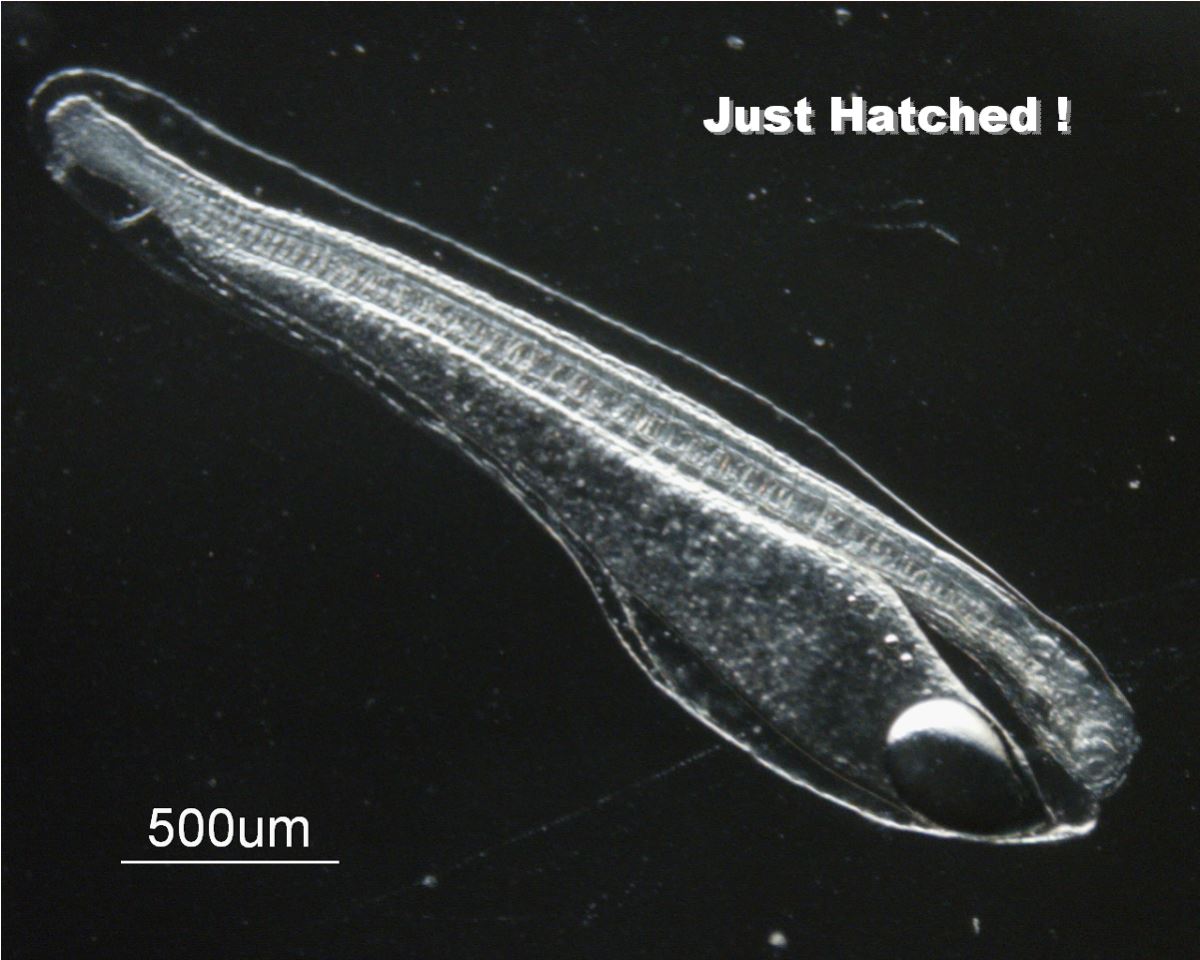
圖1.3.4剛孵化的日本鰻(Tsukamoto
et al. 2011)
剛孵化之後的鰻魚仔魚,稱之為卵黃囊期的前期柳葉鰻(Pre-leptocephalus)。前期柳葉鰻,體長約6mm,尚保有來自母體的卵黃物質而不需仰賴外界食物(圖1.3.4)。一星期後卵黃吸收殆盡,必需很快找到餌料,否則就會大量死亡,這時候是魚類生死存亡的關鍵時刻而稱之為危險期(Critical period)。為獲得豐富的餌料,前期柳葉鰻會把自己在海洋中的垂直分布調整在餌料生物(葉綠素a)分布最豐富的地方。也就是水溫躍層的頂端、鹽度極大值的深度(圖1.3.5)。2005年6月的新月晚上,日本的研究團隊在西馬里亞納海嶺南部的Suruga海山西方海域採獲130尾體長5mm左右剛孵化後兩天的前期柳葉鰻,從海流流速和孵化後經過的日數來推算,產卵場應該位於Suruga海山附近的鹽度鋒位置(Tsukamoto 2006)。這一次是繼1991年之後更精準地接近日本鰻的產卵場。
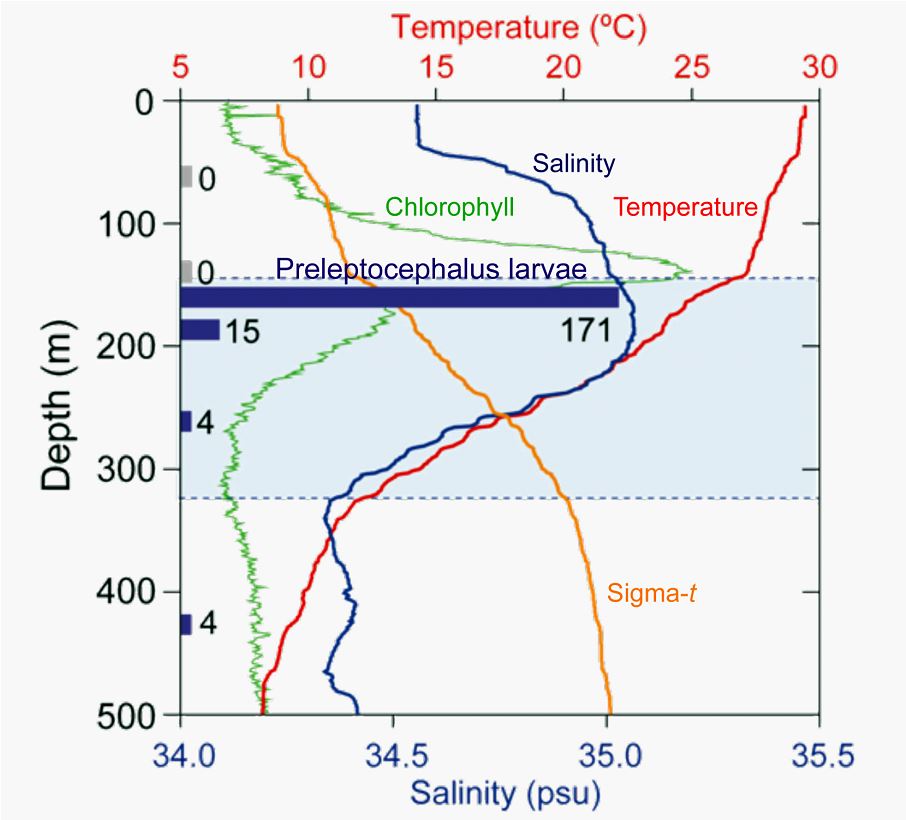
圖1.3.5前期柳葉鰻的垂直分布與葉綠素Chlorophyll、水溫、鹽度及密度Sigma-t之關係(Tsukamoto
et al. 2011)。
前期柳葉鰻有著與身體非常不搭調的針狀長齒,這樣特化的牙齒,讓人們以為前期柳葉鰻可能是肉食性(圖1.3.6)。事實上不然,該長齒並不適合捕捉會動的餌料,因為前齒突出,捕到的獵物會逃掉,這一點與長齒向內彎的鯊魚以及其他肉食性魚類不同。隨著成長,前期柳葉魚的牙齒會再吸收,與其說是用來捕食獵物,還不如說是用來儲存鈣質,提供後續的發育過程所需。另有一說法,牙齒是用來捕捉海洋上層漂下來的有機碎屑(海洋雪)等食物。
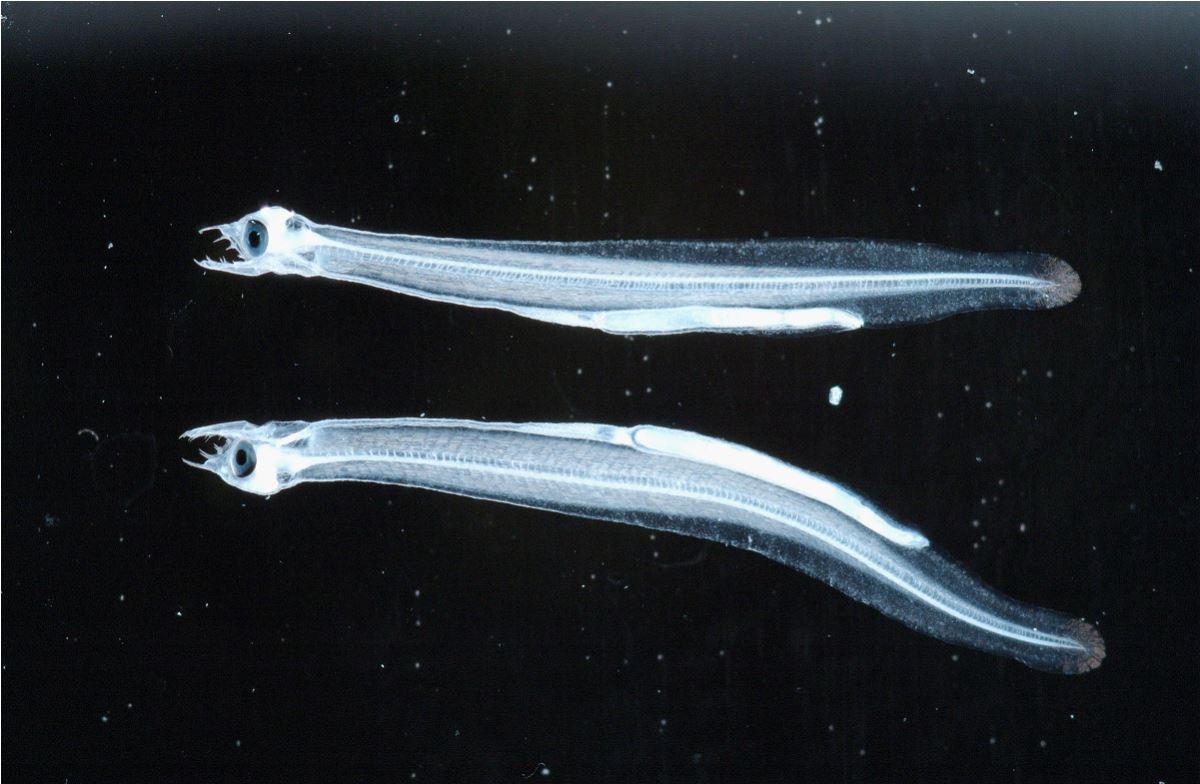
圖1.3.6牙齒特化的前期柳葉鰻(photo: Irago Institute)
1.3.2 柳葉鰻是鰻魚一生中最重要的階段(The leptocephalus is the most important life stage of eels)
柳葉鰻是魚類最特殊的發育階段。全世界2萬多種魚類中,只有海鰱目、鰻鱺目、北梭魚目以及囊腮鰻目等四個目,在早期發育階段的時候會經過柳葉魚階段。這四個目的成體外型相差極大,但DNA序列證明牠們有相近的血緣關係。在分類學上,牠們是來自共同的祖先。也因此推論淡水鰻的祖先是從深海魚類演化而來的(Inoue
et al. 2010)。前期柳葉鰻隨著成長,體高逐漸變高,形成柳葉狀,而稱之為柳葉鰻(Leptocephalus)(圖1.3.7)。柳葉鰻的體型適合其海洋浮游期(PLD)的長距離漂流。柳葉鰻的PLD長達4~11月。PLD的長短因種而異,依產卵場與陸地的距離而定,熱帶鰻較短,而溫帶鰻較長。柳葉鰻每日大約成長0.5mm,全長達60mm時就開始變態成為玻璃鰻。
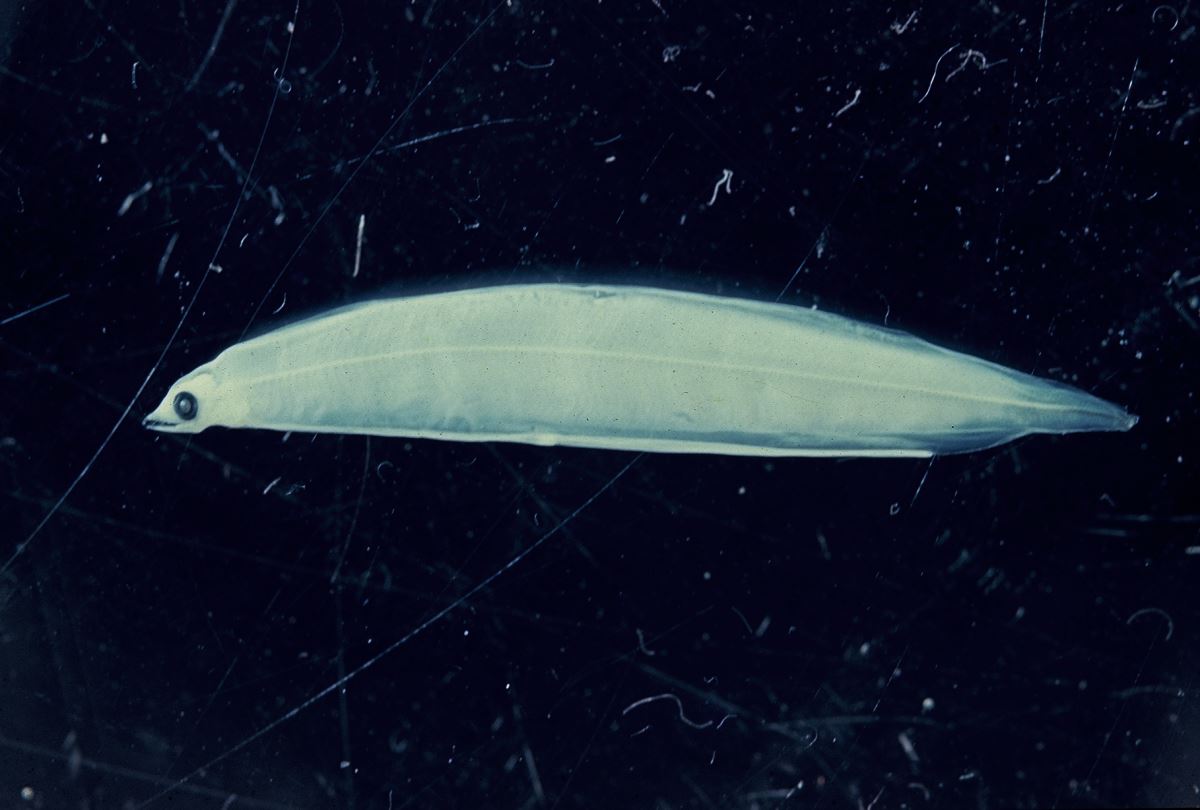
圖1.3.7臺灣的研究團隊第一次採到的日本鰻柳葉鰻標本(體長27.4mm TL)。採集時間:1995年8月22日8:43-9:14 pm,採集地點:12
o30’N,131
o30’E, 深度250m, 採集用的網具為IKMT(Liao
et al. 1996)。
(1) 浮力適應是柳葉鰻展開長距離漂游的生命旅程之關鍵(Planktonic adaptation of leptophalus for long distance dispersal)
柳葉鰻想要從大洋的產卵場長途跋涉漂游到陸地生息地的沿岸水域,必須採取最省力的漂游方式,依靠海流來輸送。然而想要隨海流漂送,首先就必須讓自己漂浮。柳葉鰻體表有大量的黏液多醣體(Glycosaminoglycan,GAG),這是一種細胞外物質,構成柳葉鰻身體很多的細胞間隙,細胞間隙蓄積的水分,可增加身體的含水量,同時體表的鹽類細胞,可幫助滲透的調節,使得GAG間隙的水分保持比外界海水低張的狀態,身體才能漂浮起來(Kaneko
et al. 2003, Tsukamoto
et al. 2009a)。隨著成長GAG的蓄積量增加,柳葉鰻的比重進一步變小,而能漂浮,才能被海流輸送到遠方(圖1.3.8)。
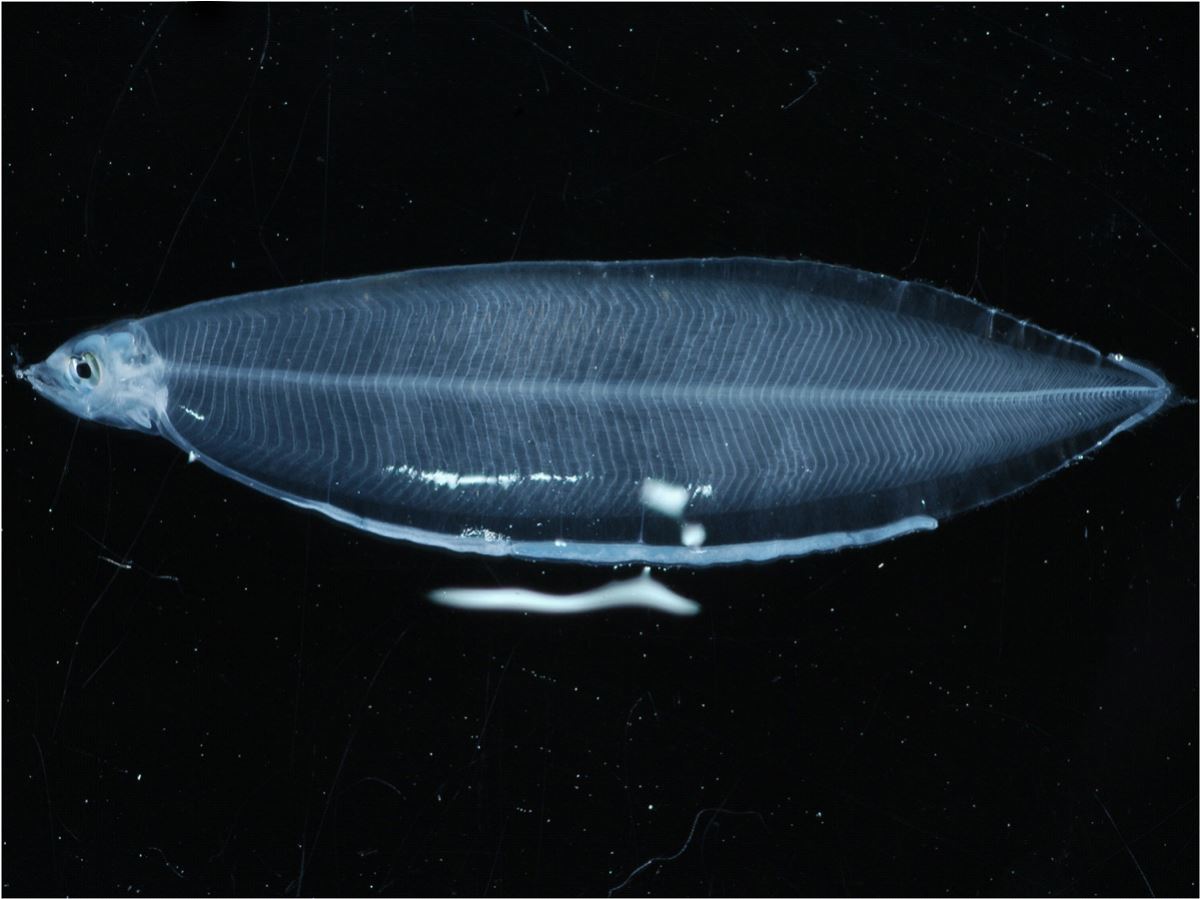
圖1.3.8柳葉鰻利用黏液多醣體調節滲透壓增加浮力(photo: Shizuoka Prefectural Fisheries Experimental Station)
(2) 柳葉鰻的變態是生命的轉捩點(Metamorphosis of leptocephalus is a key stage of life)
柳葉鰻的變態是離開黑潮強流帶進入沿岸水域的關鍵階段(圖1.3.9.1)。如果不變態的話,柳葉鰻會一直順著黑潮漂
往外洋,
而走上生命的不歸路。就像毛毛蟲變蝴蝶
、蝌蚪變青蛙一樣
,柳葉鰻長到一定體長,大約60mm,到達陸地邊緣的水域之後,身體起了很大的變化
。義大利科學家Grassi and Calandruccio(1897) 最先觀察到柳葉鰻變態為玻璃鰻的現象(圖1.3.9.2)。
柳葉鰻的變態,只需在2~3星期間,
變態時身體上的GAG急速分解,提供身體的骨骼發育及變態
所需的能量之用。
變態之後,因GAG的消失,比重變大
,。此時身體已由柳葉型變成流線型
的玻璃鰻,因此運動方式由被動性的浮游變成主動性的洄游
,而能離開黑潮強流帶進入沿岸水域。同時,體表的鹽類細胞逐漸消失,由鰓的鹽類細胞取而代之
,因此身體滲透壓調節的
能力加強。變態時身體有數mm的縮小(負成長),
變態後食性改變,開始快速成長。變態不只
是形態變化,生理機能和生態特性也激烈變化。

圖1.3.9.1柳葉鰻的變態。身體由柳葉型變成流線型玻璃鰻,運動方式也由被動性的浮游變成主動性的洄游(photo: Hideki Tanaka)

圖1.3.9.2義大利科學家Grassi and Calandruccio 最先觀察到柳葉鰻變態為玻璃鰻的現象(photo: Mari Kuroki)
(3) 柳葉鰻的旅途終點站(End of larval migration)
柳葉鰻的變態個體,至今只捕獲6尾。變態期只有短短3個星期左右,而且分布及行為都有所變化,採集不易(Shinoda
et al. 2011)。六尾變態中的柳葉鰻,多數採自台灣東方黑潮渦流域,黑潮的沿岸側採到的都是變態後的玻璃鰻。在黑潮中變態完成後,柳葉鰻即脫離黑潮進入沿岸域。變態日齡是決定玻璃鰻進入那個國家的時間點。台灣與日本的柳葉鰻的變態時間相差3個星期左右,早變態者進入台灣,晚變態者則漂到日本(Cheng and Tzeng 1996, Kuroki
et al. 2008a)。
變態時間決定旅途終點的道理,亦可用來解釋美洲鰻與歐洲鰻的分離洄游機制(圖1.2.1)。這兩種大西洋鰻都誕生在大西洋的百慕達神秘三角洲的藻海,之後進入北赤道洋流,由東向西流,然後進入灣流(Gulf Stream),沿美洲東岸洄游,美洲鰻先變態進入北美洲的河川流域,而歐洲鰻則要橫越北大西洋,到達歐洲國家的沿岸水域後才變態。歐洲鰻利用延遲變態來延長柳葉鰻階段,以便到達較遠的歐洲國家(Wang and Tzeng 1998,2000)。
1.3.3玻璃鰻溯河過程中的色素發育(Pigmentation of glass eels during upstream migration)
(1)體表色素隨發育階段的變化
從柳葉鰻到玻璃鰻乃至鰻線的色素發育,可分成I-VI六個階段(Bertin 1956,Strubbery 1913)。I-IV是柳葉鰻從完全成長之後到變態完成為止的階段(圖1.3.10)。柳葉鰻全身透明,體型側扁、狀似柳葉,完全是為了適應外洋受海流攜帶的被動洄游。柳葉鰻長到最大體長(日本鰻大約60 mm)之後,變態為玻璃鰻。變態時體長會縮小(Bertin 1956的Fig.38)。
玻璃鰻階段,脊椎骨很快發育完成、長出肌肉,體型變成強而有力流線型,主動洄游離開黑潮,由海水游向淡水進入河川。玻璃鰻向岸洄游的過程中,皮下色素開始發育形成保護色。V
A到VI
A4是玻璃鰻至鰻線的皮下色素發育階段(圖1.3.10)。
.jpg)
.jpg)
.jpg)
VA VB VA
.jpg)
.jpg)
VI
A1 VI
A2
.jpg)
.jpg)
VI
A3 VI
A4
圖1.3.10玻璃鰻到鰻線的發育過程中軀幹部皮下色素分佈之變化。VA階段: 頭頂及軀幹皆無色素。VB階段: 只有頭頂出現色素。從VI
A1階段至VI
A4階段背鰭,其基底至腹部逐漸出現色素(Strubbery 1913)(photo:何敏和韓玉山)
(2)體表色素分佈的種類間之差異
臺灣河口域常見的玻璃鰻種類,有日本鰻、鱸鰻、和太平洋雙色鰻,其身體比例和尾部黑色素胞互異(圖1.3.11a,b)。日本鰻和鱸鰻,其背鰭起點和肛門之間的脊椎骨數分別為8-11節和13-17節,故稱之為長鰭鰻。而太平洋雙色鰻,其背鰭的起點和肛門之間的脊椎骨數為0-2節,故稱之為短鰭鰻(曾1983)。日本鰻玻璃鰻的體長範圍為54-58mm,鱸鰻為46-52mm,太平洋雙色鰻為44-50mm(曾1983)。
剛剛到達沿岸的玻璃鰻,其色素發育階段為VA,頭頂及軀幹皆無色素,但尾部開始出現色素而且因種而異。漁民在河口捕撈玻璃鰻時,就依黑色素胞的出現與否及其出現位置來區別鰻苗的種類。全身透明無黑色素胞的日本鰻,漁民將之稱為白鰻。尾鰭出現黑色素胞的太平洋雙色鰻和尾柄部出現黑色素胞的鱸鰻、西里伯斯鰻和呂宋鰻,統稱之為黑鰻。利用色素胞的有無來挑選出日本鰻。過去,日本鰻是台灣唯一養殖的種類。但近年來日本鰻鰻苗缺乏,同時2009年4月農林廳解除鱸鰻保育禁令,漁民也開始養殖鱸鰻。

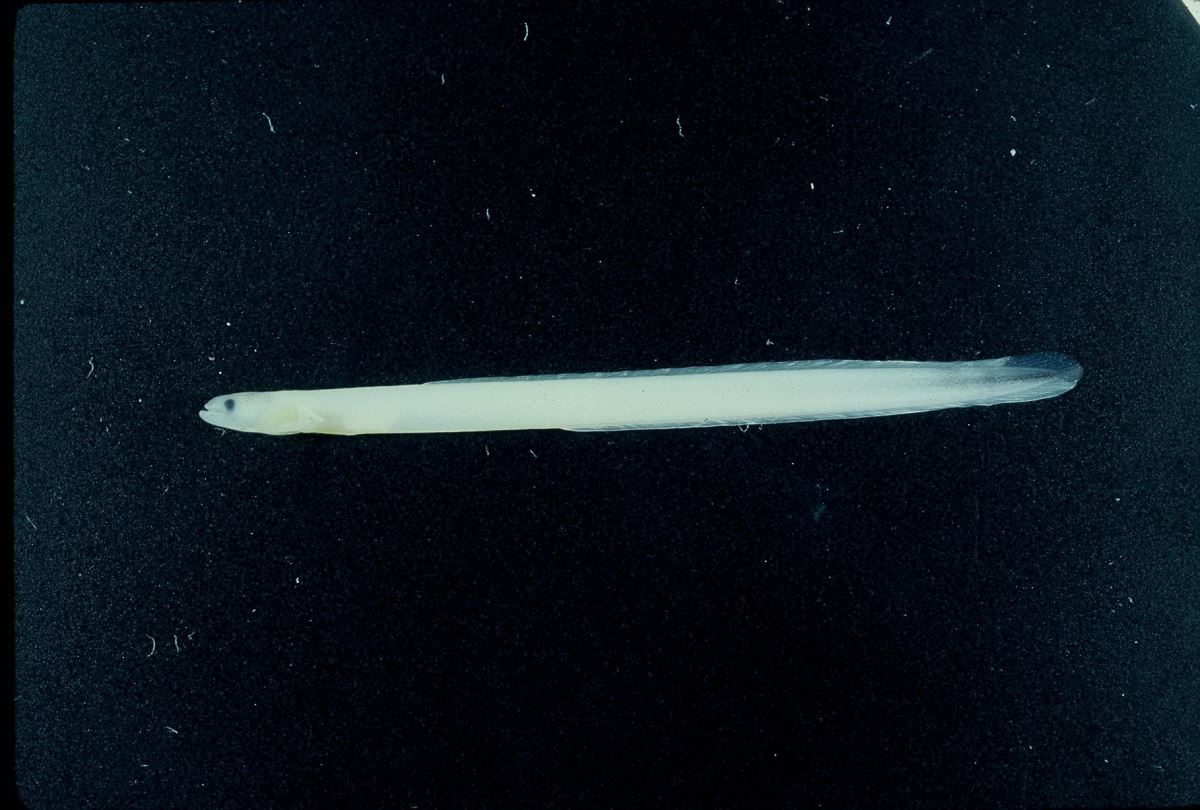
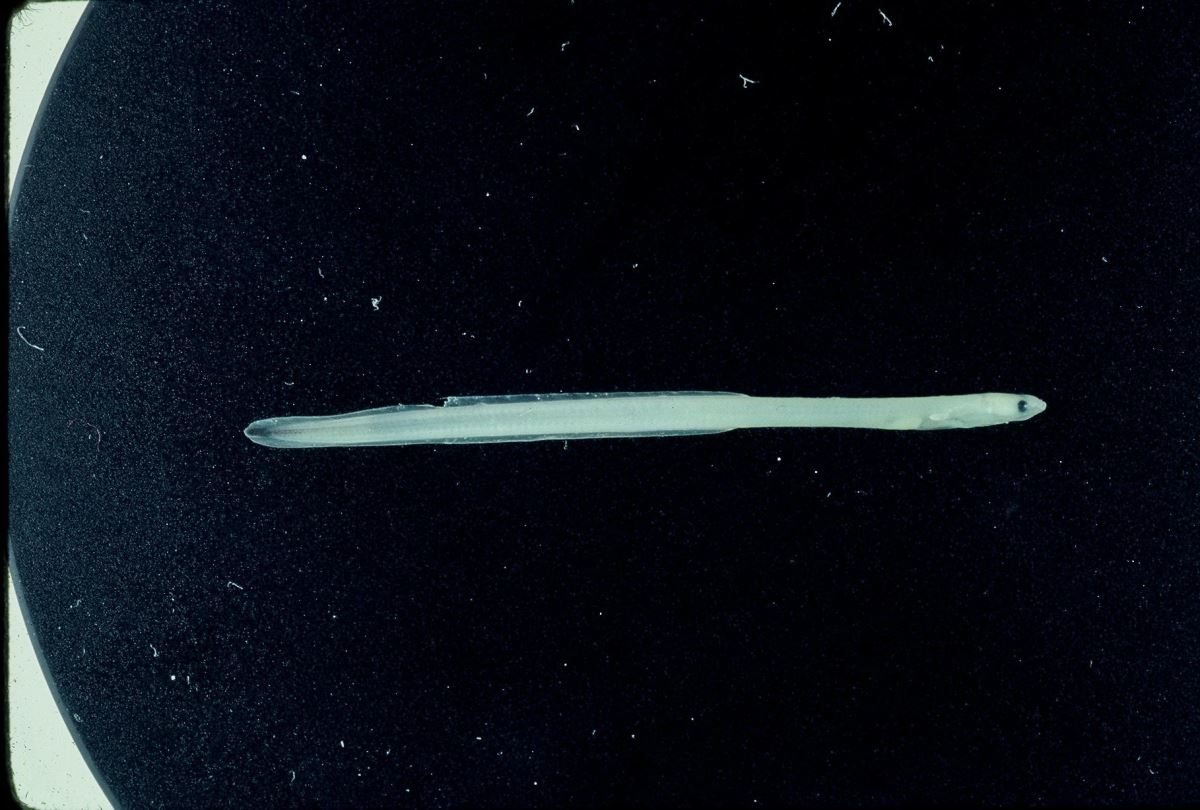
圖1.3.11a玻璃鰻的外觀。(上)日本鰻;(中)鱸鰻;(下)太平洋雙色鰻
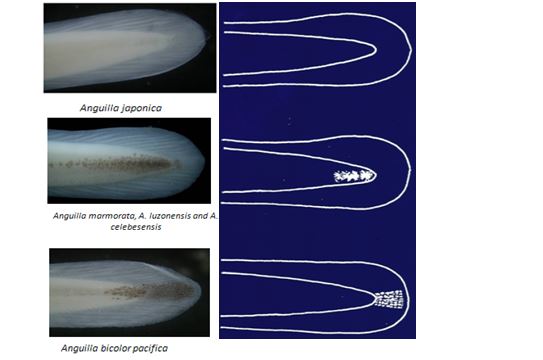
圖1.3.11b玻璃鰻尾柄部和尾鰭的黑色素胞出現情形及示意圖。(上)日本鰻尾柄部和尾鰭皆無黑色素胞;(中)鱸鰻、呂宋鰻和西里伯斯鰻的尾柄部有黑色素胞;(下) 太平洋雙色鰻的黑色素胞出現在尾鰭
根據Tzeng(1985)於1981~1984年在台北縣福隆雙溪沿岸的調查,剛剛到達沿岸的玻璃鰻,其大部分的個體之色素發育階段為VA(佔調查樣本數1185尾的93.7%),其次是VB(5.2%)和VI
A1(1.1%)。進入河川之後皮下色素逐漸增加,VB階段的個體之比例逐漸增加,VA下降到18.9%。從VB的 玻璃鰻階段到VI
A2的鰻線階段,體長會縮小5%左右。VI
A3以後,體長逐漸增大。
玻璃鰻的日文叫シラス(白子)ウナギ。鰻線的日文叫クロコ(黑子)。兩者的界線不是很明確, 一般認為VI
A2為止的階段叫玻璃鰻,VI
A3以後叫鰻線(Fukuda 2010)。
1.3.4玻璃鰻的向岸洄游生態(Ecology of the estuarine migration of glass eels)
鰻線到達台灣河口時的全長,日本鰻約54.0~58.0mm,比熱帶性鱸鰻的46.0~52.0mm稍大(曾 1983)。分布在亞寒帶的歐洲鰻,其鰻線之全長可達75mm(Jesperson 1942)。鰻線到達河口域時候的日齡,因分布域的緯度不同而有所變異,熱帶性的種類約3~4個月大,日本鰻大約為5~6個月大(Tzeng 1990,Tzeng and Tsai 1992,Cheng and Tzeng 1996)。美洲鰻鰻線到達河口的日齡與日本鰻相似,大約是5~6個月。歐洲鰻則要1年以上才能從產卵場到達河口(Lecomte- Finiger 1992,Wang and Tzeng 1998,2000)。鰻線到達河口域時候的日齡,可利用耳石上的日周輪來推算
。玻璃鰻進入河口後,喜歡躲藏在落葉或泥巴中,以便攝取食物和逃避敵害(圖1.3.12)。


圖1.3.12 (上圖)玻璃鰻活體全身透明,(下圖)連脊椎骨和內臟都看得很清楚。黃色部分為水筆仔(紅樹林)的漂流落葉(photo:廖運志)
鰻線靠岸的季節及持續時間,因種而異,溫帶鰻集中在冬季,而熱帶性鰻幾乎全年出現。鰻線靠岸的習性也因種而異,因此台灣東西岸鰻線的種類組成有很明顯的差異性(圖1.3.
1413)。日本鰻的玻璃鰻是隨著中國大陸沿岸流到達台灣西部沿岸的,而鱸鰻的玻璃鰻則是隨著黑潮與黑潮支流到達台灣東部和西南部沿岸的。因此,北部的淡水河以日本鰻 (
A. japonica) 為主,東部的秀姑巒溪和南部的枋山溪以鱸鰻(
A. marmorata)為主。台灣西南海域東港溪、東北部宜蘭河是中國大陸沿岸流和黑潮的折衝地帶,日本鰻和鱸鰻平分秋色。這樣的分配與鰻線上溯時,對溫度的偏好和海流輸送有關(Han
et al. 2012)。鱸鰻是熱帶性品種,在高水溫時才會上溯河川,受黑潮影響的東部水溫高,來游量自然比較多。太平洋雙色鰻(
A. bicolor pacifica)和呂宋鰻(
A. luzonensis) 是台灣以南的品種,相對數量較少。
鱸鰻分布於低緯度地區(菲律賓、印尼)之數量遠高於高緯度地區(日本)。鱸鰻終年皆可產卵,因此其鰻苗終年皆可捕獲,在臺灣主產期在每年3-10月。太平洋雙色鰻屬於熱帶品種,廣泛分布於菲律賓與印尼,台灣為其分布北界,因此數量很少,其鰻苗終年皆可捕獲,在臺灣主產期在每年秋冬。近來在台灣與菲律賓呂宋島發現的新種呂宋鰻對其資源分布與生活史所知不多,在臺灣主產期在每年秋季,數量很少。
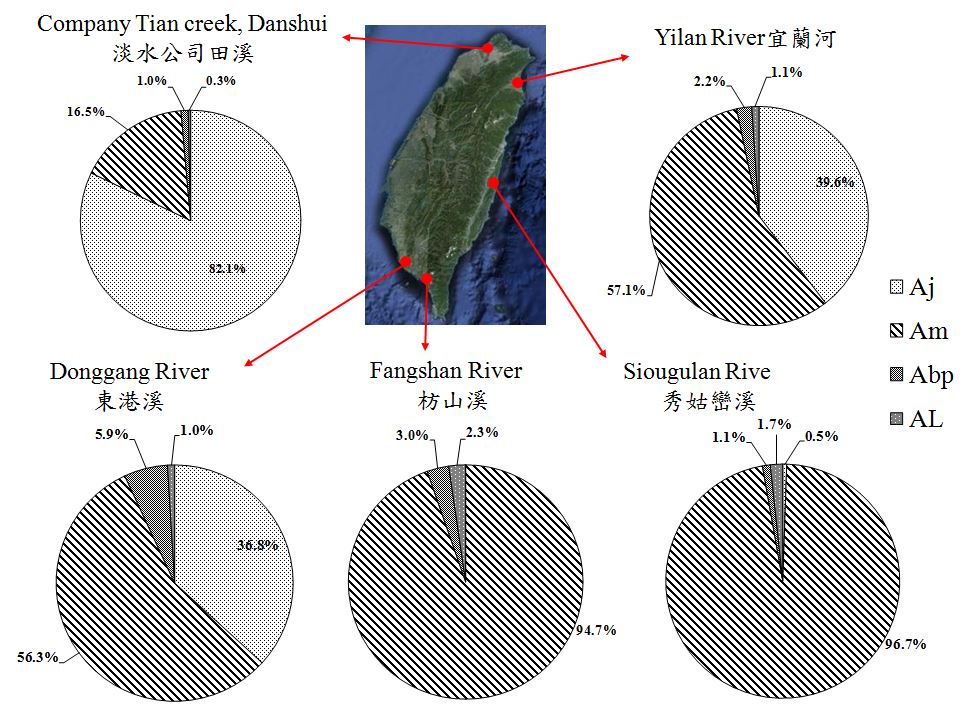
圖1.3.
13灣沿岸玻璃鰻種類組成的變化
日本鰻是臺灣水產養殖的重要魚種。台灣地區每年的日本鰻鰻苗汛期約始於10月下旬, 3月下旬結束,12月到2月是台灣日本鰻鰻線的盛產期,西岸各縣市皆出現鰻線。整體而言,台灣沿岸各縣市日本鰻鰻線,以宜蘭縣的產量為最多,持續月份也最長(圖1.3.
14)。
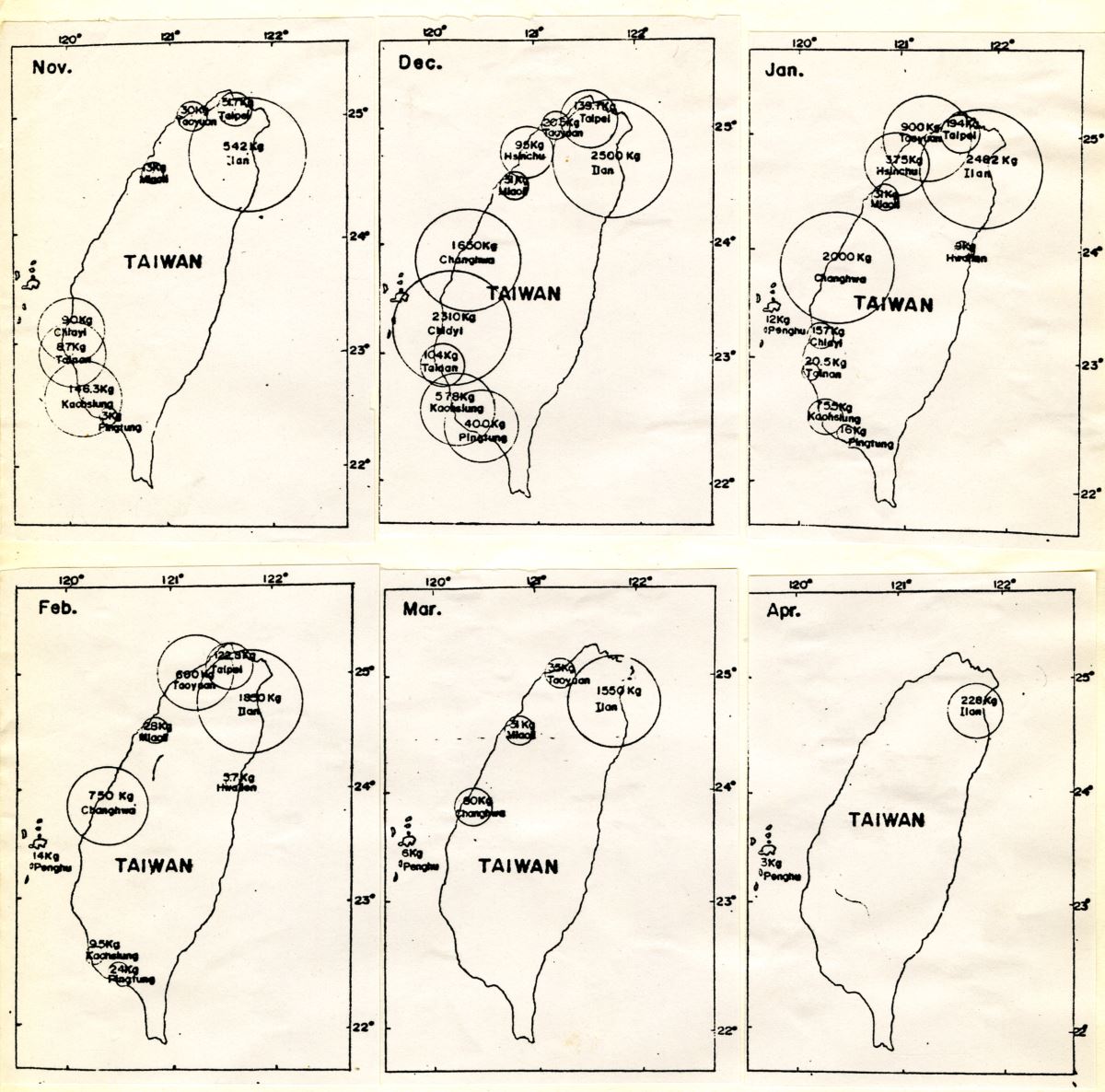
圖1.3.
14台灣沿岸各縣市1970-1971年日本鰻鰻線產量的月別變化(引自陳宗雄 1975)
宜蘭是日本鰻鰻苗洄游必經之地,台灣鰻苗生產的第一重鎮。因地處冬季黑潮水與沿岸水交匯的地區,有利於鰻線的聚集。其平均年產量佔全島4成以上,對臺灣鰻魚養殖業貢獻非常大。每到鰻苗的捕撈季節,除了宜蘭當地的漁民外,很多來自花東的原住民,在蘭陽溪口兩岸的沙灘上,搭起了一座座的帳篷,作為臨時放置捕鰻器具之所,原住民更以帳篷為家,在此度過長達4個月的捕苗季,賺取生活費,補貼家計。
1.3.5成長期的黃鰻(The yellow eel)
鰻線溯河之後,變成黃鰻。黃鰻是鰻魚的成長和定棲的階段。身體的顏色會隨著棲地環境的顏色改變,養殖的鰻魚,有黑色、青藍色和黃綠色等(圖1.3.
15)。

圖1.3.
15黃鰻體色會隨環境而改變
鰻魚的成長率,可利用耳石上的年輪的年輪半徑與捕獲時的魚體長來逆算每一個年輪形成時的體長,然後可求得其成長曲線。以2005年~2007年高屏溪野生的日本鰻為例,第一年大約可長到20cm左右,第三年35cm,第五年將近50cm,第七年70cm左右(Tzeng
et al. 2000, Lin
et al. 2007)。
1.3.6 黃鰻的巢穴、領域及歸巢行為(Nest, home range and homig of yellow eels)
進入定居生活的黃鰻,其住家有多種形式,包括岩石遮蔽的陰暗處,石塊下方,枯木下方,泥中的巢穴以及草叢等等。能夠滿足其背光性和受干擾後能逃避的條件即可。黃鰻原則上行單獨生活,歐洲鰻和美洲鰻在越冬時有成群的習性(LaBar
et al. 1983,1987)。一般而言,小型的黃鰻生活在淺水區,大型者則生活在較深的場所(Yokouchi 2009)。但有時在淺的地方也會發現大的個體,嚴格來說大小分棲的情形,並沒有那麼明顯。鰻魚喜歡在柔軟泥中築巢穴。
生物的覓食及生活圈範圍稱之為歸巢性。這包括離巢及索餌的範圍。定居期的黃鰻,也有歸巢行為。在日本西神田川的日本鰻的標識放流調查中,發現3年的移動範圍只有710m (Yokouchi
et al. 2009),而2002~2006年在台灣南部的高屏溪所標識2473尾黃鰻的最大移動範圍也都在1km以內(Lin
et al. 2012a)。由此可見,鰻魚在河川中的領域範圍並不大。
美國密西西比河下游的美洲鰻和荷蘭Elbe河的歐洲鰻的研究案例,也顯示其移動距離大部分都在放流點幾十公尺以內。在湖內利用無線電波追蹤其行為,也發現其活動範圍在2公頃以內,這些顯示黃鰻的定棲性很強。另一方面發現越冬時,黃鰻會把在河川的移動範圍轉移到沿岸域,相反地,季節交換後會再回來,表示環境條件苛刻時,會改變棲地。
鰻魚會不會像鮭魚或鴿子一樣有歸巢的行為,利用超音波標識實驗,把美洲鰻放流到離採集點10~17km遠的河川上游或下游後,發現56%的標識鰻在9天之內會回到原點(Parker 1995)。這表示鰻魚可以感知潮流及利用嗅覺的本能做定向洄游回到家(Barbin 1998)。在荷蘭的調查也發現,把110尾歐洲鰻放到100公里以外的外海有20尾會回到原來的地方(Deelder and Tesch 1970),顯示鰻魚可能有感知地磁的能力,在很短的時間會回歸到原來的巢穴。
1.3.7 環境決定鰻魚的性別(The environmental sex-determination hypothesis for the eel )
(1)天然鰻:雌多於雄
鰻魚的性別決定屬於後天型,當幼鰻成長至15-30公分後,生殖腺才開始發育,一直到降海洄游前都處於低度發育的階段,無怪乎古人認為鰻魚是''無性生殖'',仔魚是從泥巴中蹦出來的。台灣的天然河川中鰻魚雌雄的比例非常懸殊,雌魚佔80%以上。為何如此呢? 一般來說雌魚的體型愈大,孕卵數越多,產生的後代也愈多。而一隻雄鰻魚可以配數隻雌鰻魚,因此當雌鰻比例較多時,可產生較多子代,有利於族群的增大。控制鰻魚性別的外在原因不明,可能與族群密度、成長速率、棲地鹽度等等有關。
(2)養殖鰻:雄多於雌
在養殖環境中,神奇的事情發生了,雌雄的比例同樣非常懸殊,但卻是雄魚佔90%以上,和野生的情形剛好相反。雄鰻的生殖腺通常比雌鰻的要小很多,不仔細看常常難以發現。因此大部分的養殖漁民都不知道原來鰻魚有分公母。雄鰻的體型也比雌鰻小一號,若以一般上市規格 200-250克/公斤,雌雄鰻魚皆可輕易達到此體型,但是如果體型在400克以上則多半是母鰻了。
(3)高屏溪日本鰻的陰盛陽衰現象
高屏溪是台灣南部最大的河川。高屏溪下游是日本鰻的重要棲息地,野生鰻魚很多。
從1998年開始臺灣大學漁業科學研究所曾萬年教授的研究團隊每年調查高屏溪日本鰻的性別比例時,發現100條鰻魚中有80條都是雌鰻,可是到了2004年7月~10月這段時間,卻發現雄鰻的比例突然增加,雌鰻的比例下降到20%左右。同年的11月後逐漸恢復到2004年之前的雌鰻比例多的現象(圖1.3.
1716)。
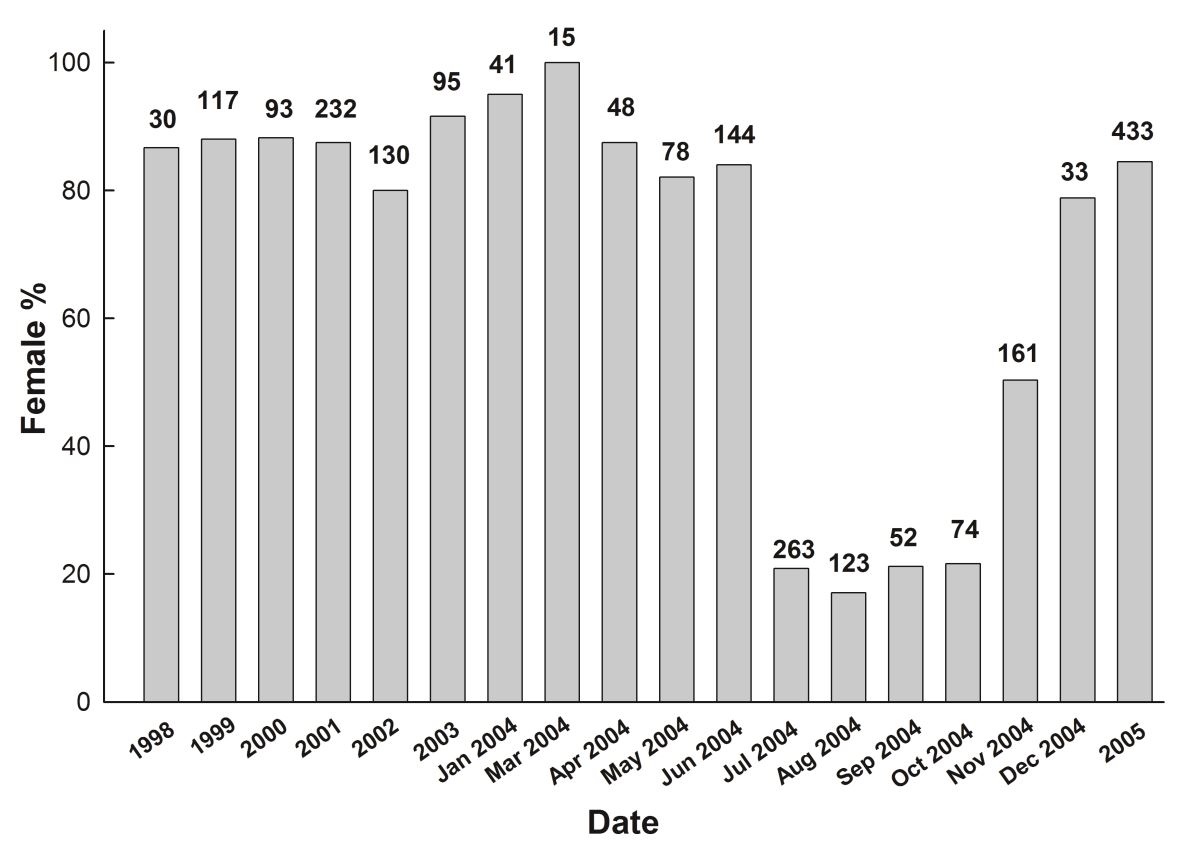
圖1.3.
1716高屏溪日本鰻族群的雌魚比例之時間變化(圖中數字是樣本數)
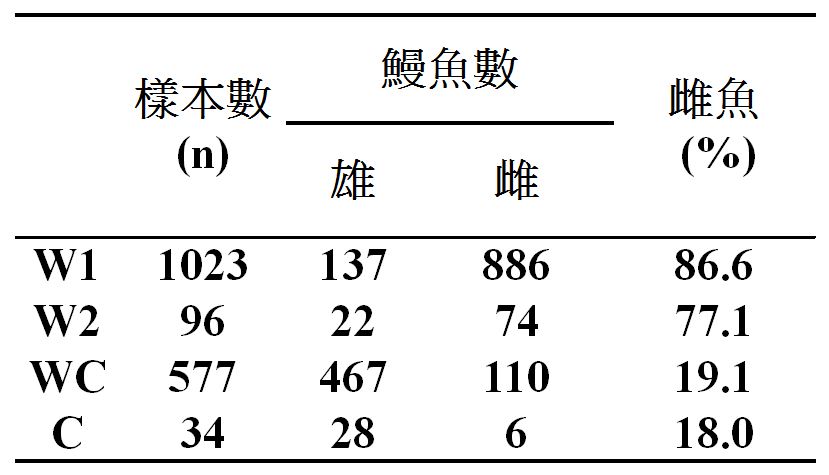
高屏溪日本鰻的性別比例變化之原因,原來是敏都利颱風來襲,造成高屏溪附近的養殖池潰堤,大量的養殖鰻從養殖池逃到高屏溪所致(Chu et al. 2006)。這些養殖鰻不到4個月就被漁民捕光了,因此很快地高屏溪的天然鰻又恢復到雌多於雄的現象。由表一的調查統計數字來看,得知2004年之前(W1)和2004年7~11月(W2) 野生日本鰻的雌鰻比例分別為86.6%和77.1% 。可是,逃到高屏溪的養殖鰻(WC)的雌鰻比例為19.1% 與在養殖池的雌鰻(C)比例18.0%非常接近。這證明天然鰻族群中的性別比例為雌多於雄,而養殖鰻則雄多於雌。
表一
天然鰻(W) 與逃到高屏溪的養殖鰻(WC)和養殖池的養殖鰻(C)的雌魚比例之比較
(4)族群密度與鰻魚的性別決定機制
鰻魚一生下來就沒有性染色體,無法辨別雌雄性別。牠的性別非先天遺傳所決定,而是後天環境所使然。決定鰻魚性別的環境因子,眾說紛紜,例如族群密度、成長速率及棲地鹽度等都有關係。其中一項比較受重視的是族群密度。從我們的調查中,發現族群密度愈低則魚群中雌魚所佔的性別比例愈高;反之,隨著魚群密度的增加,則雌魚比例逐漸降低(Han and Tzeng 2006)。
鰻魚性別的決定與其族群密度的關係,可以用族群成長的邏輯曲線來加以解釋(Colombo and Rossi 1987)(圖1.3.
17)。玻璃鰻剛進入河川時,性別尚未分化,成長到黃鰻階段後,少部分自然分化為雌或雄。但大部分中間性別的黃鰻要分化為雌或雄,則視環境所容許的族群最大負載量(K)而定。在環境負載量低時,中性的黃鰻分化為雌性,並進一步成為銀鰻;反之,若族群密度超過負載量時則分化為雄性銀鰻。鰻魚之所以做這樣的性別選擇,乃受限於環境資源。為了達到共存共容,繁殖更多子代,鰻魚採取兩性分工的生存策略,也就是雄魚性成熟年齡最小化(Age minimization),雌魚成熟體長最大化(Size maximization)的生存策略。因為河川環境變數太多,而且每年從外海加進來的新生代不確定性也很高。族群密度高時食物不夠分配,分化為雄性,提早成熟,早日離開擁擠的河川棲地,降海進行生殖洄游,可疏解族群壓力。反之當資源密度低時,每一尾鰻魚可以分配到的食物比較多,變為雌魚,晚熟成長到較大的體長,可增加孕卵量產生更多的子代,以利族群量快速恢復,鰻魚性別選擇可以說已經到了生存策略演化的最高境界。
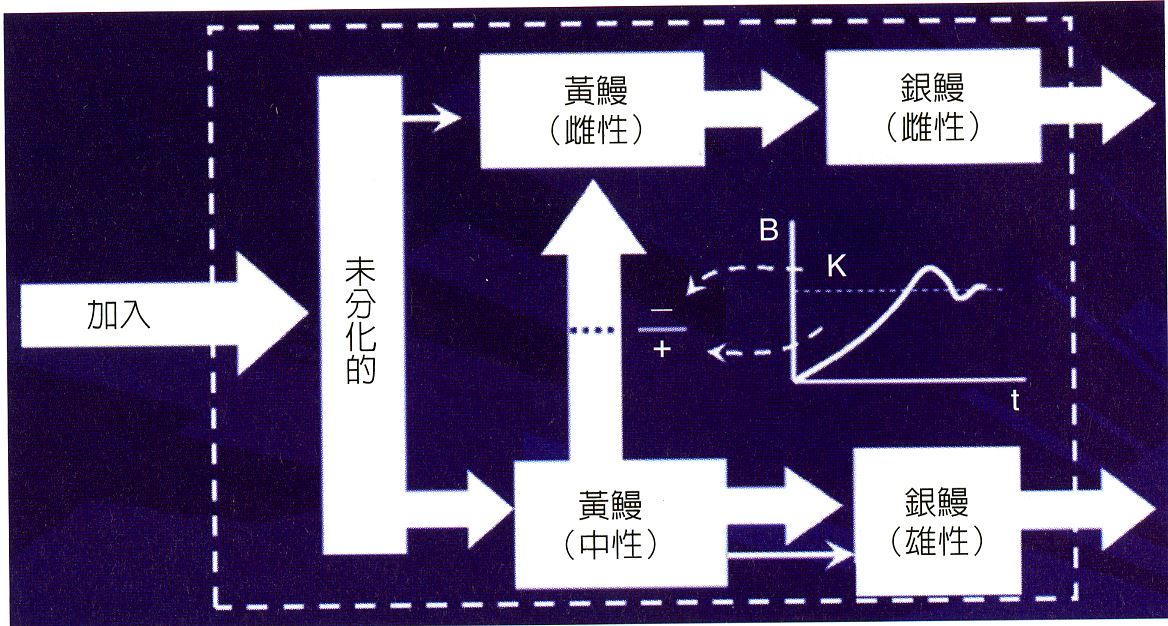
圖1.3.
1817鰻魚的性別決定機制。內插圖是族群成長的邏輯曲線,當族群量(B)很低時鰻魚傾向分化為雌性;反之,當族群量(B)超過環境負載量(K)時則分化為雄性。
1.3.8 鰻魚的壽命(Eel mean life span)
鰻魚是壽命比較長的魚類。鰻魚一生只生殖一次,生殖後就使死亡,性成熟年齡就是牠的最大壽命。鰻魚的種類不同,壽命也不一樣。歐洲鰻的雌、雄平均成熟年齡分別為10歲與7.5歲。 美洲鰻的平均成熟年齡,雌、雄鰻分別為19.3歲與12.7歲。中國珠江產的日本鰻,雌、雄平均成熟年齡則分別為8.3歲與6.4歲。台灣產的日本鰻,雌、雄平均成熟年齡則分別為6.7歲與5.9歲。一般來說,生活在越高緯度的鰻魚,因成長速度較慢,因此成熟年齡也較晚。而有紀錄的鰻魚最大壽命,例如日本濱名湖曾捕獲一尾雌日本鰻22歲(Sudo 2011),愛爾蘭的歐洲鰻雄33歲,雌57歲(Povle & Reynolds 1996), 紐西蘭大鰻
A. diffenbachii,35歲(Todd 1980)。亦有傳說紐西蘭大鰻有超過100歲者。
1.3.9 銀鰻(The silver eel)
銀鰻是鰻魚發育和成長過程的最後階段。銀鰻在降海產卵前,消化道開始萎縮不再進食。產卵洄游及卵巢發育所需的能量,全靠降海產卵前在陸地河川攝食所累積的脂肪來供應。降海洄游之前,眼睛會變大、背部和胸鰭變黑、腹部變銀灰色,以便適應產卵洄游的深海環境(圖1.3.
2118)。產完卵完成傳宗接代的任務後,就葬身海底結束其生命。圖1.3.
2219是日本水產廳的研究人員於2009年6月22 和23日在馬里亞納島西側的海域日本鰻產卵場採獲的雄銀鰻和雌銀鰻。由此,證明日本鰻在馬里亞納島西側海域產卵。
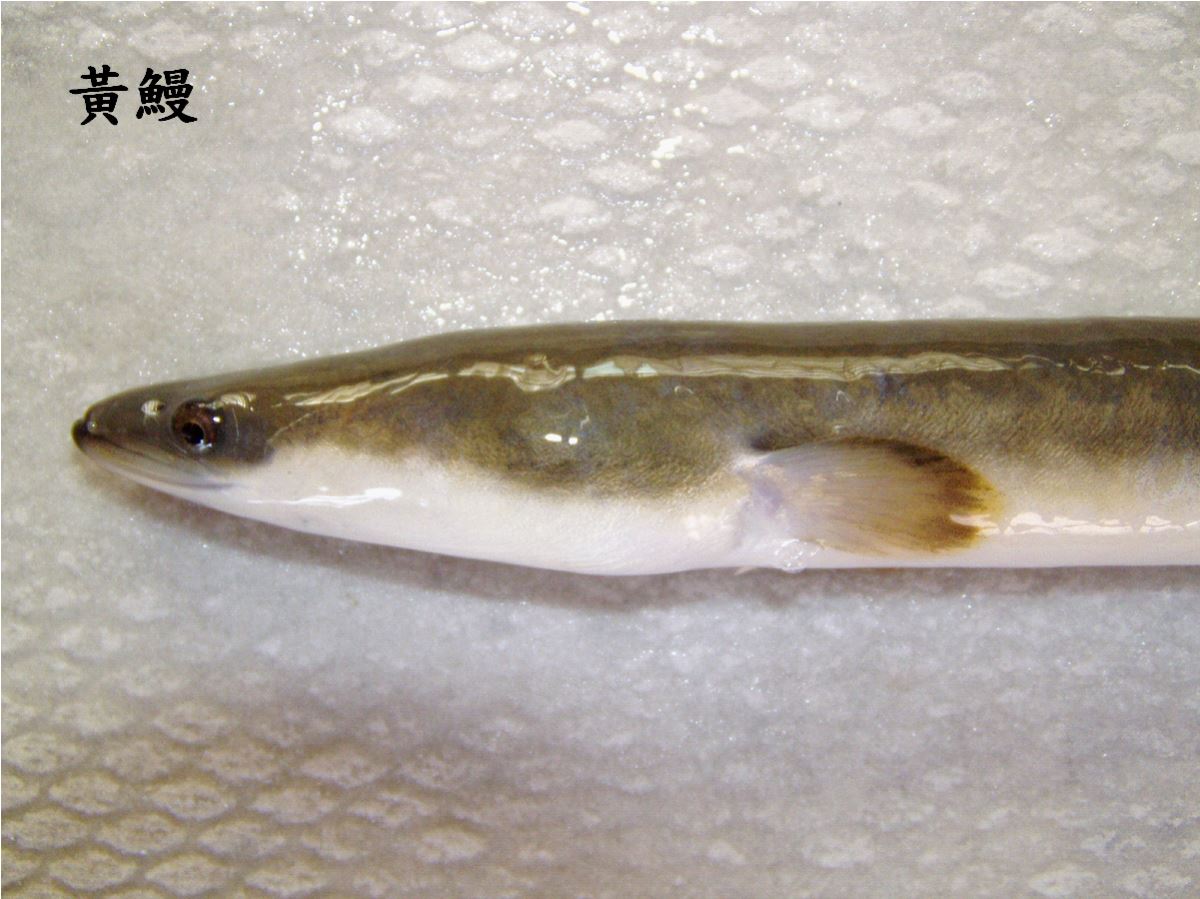

圖1.3.
1918日本鰻降海產卵前從黃鰻(上圖)變態為銀鰻(下圖)之外觀變化 (photo:Yu-San Han)
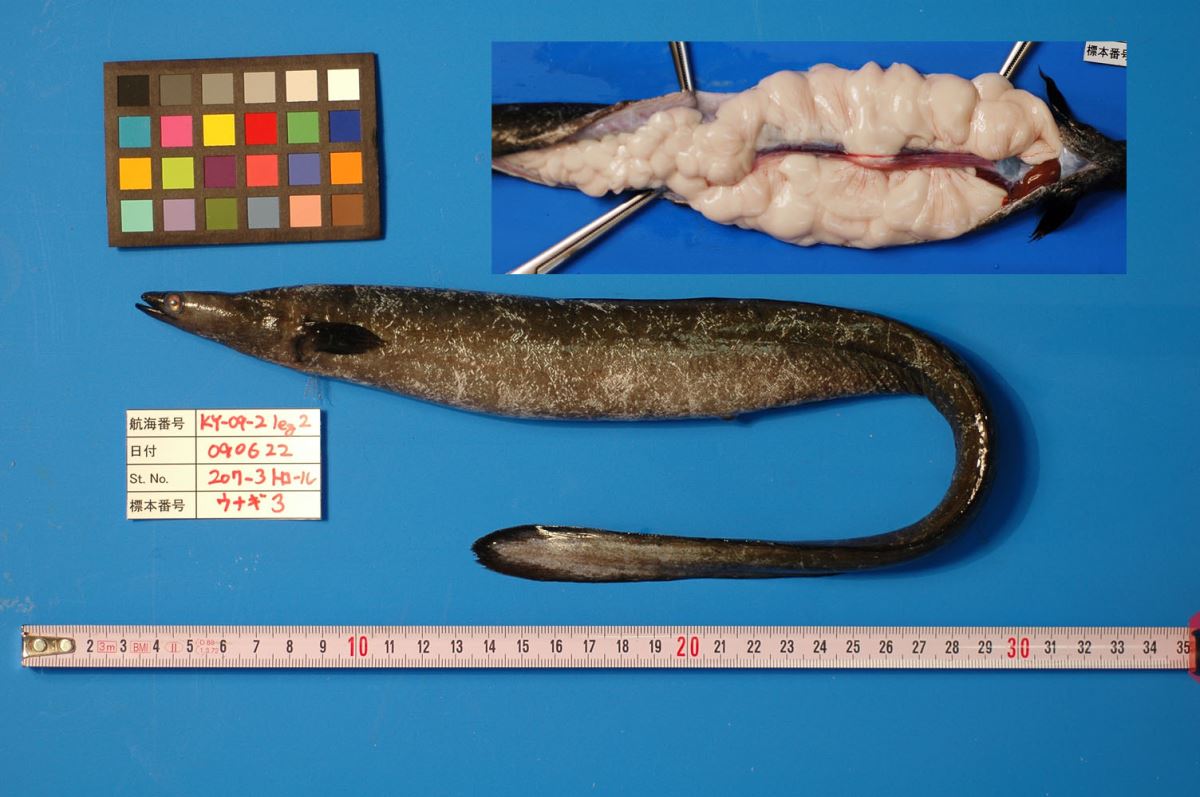

圖1.3.
19 日本水產廳的研究人員於2009年6月22 和23日分別在12-19'N, 141-35'E 和 12-17'N, 141-55'E的馬里亞納島西側海域日本鰻產卵場採獲兩尾日本鰻銀鰻。(上圖)雄性銀鰻: 體長44.7 cm TL, 體重129 g, 生殖腺重52 g。 (下圖)雌性銀鰻: 體長74.9 cm TL, 體重244 g, 排出的液體中含有卵(Photo: Hiroaki Kurogi, copyright: Fisheries Agency, Japan)
1.3.10 日本鰻的族群遺傳結構(Population genetic structure of Japanese eel)
歐洲鰻(
A. anguilla)是最早開始族群遺傳結構研究之淡水鰻 (
Ligny and Pantelou 1973)。粒線體 DNA 的研究結果,顯示歐洲鰻沒有明顯的族群遺傳分化現象(Dannewitz
et al. 2005)。日本鰻的族群遺傳結構之相關研究起步較晚,利用粒線體DNA或微衛星DNA分析日本、台灣、中國大陸等地的日本鰻鰻線標本,也發現無論在空間上或時間上,其族群遺傳結構皆無明顯分化現象( Han 2012;Han
et al. 2008,2010;Ishikawa
et al. 2001;Sang
et al. 1994)。換言之,西北太平洋的日本鰻應屬於單一逢機交配族群。
一般來說,物種的棲地範圍越大,就越容易產生地理上的隔離之族群分化現象。日本鰻的地理分怖範圍雖然大,卻沒有族群分化現象。這是因為日本鰻有一個特定的產卵場,不管子代擴散範圍有多廣,親魚最終都會回到出生的產卵場繁殖下一代,因此不易造成族群分化現象(圖1.3.
2320)。
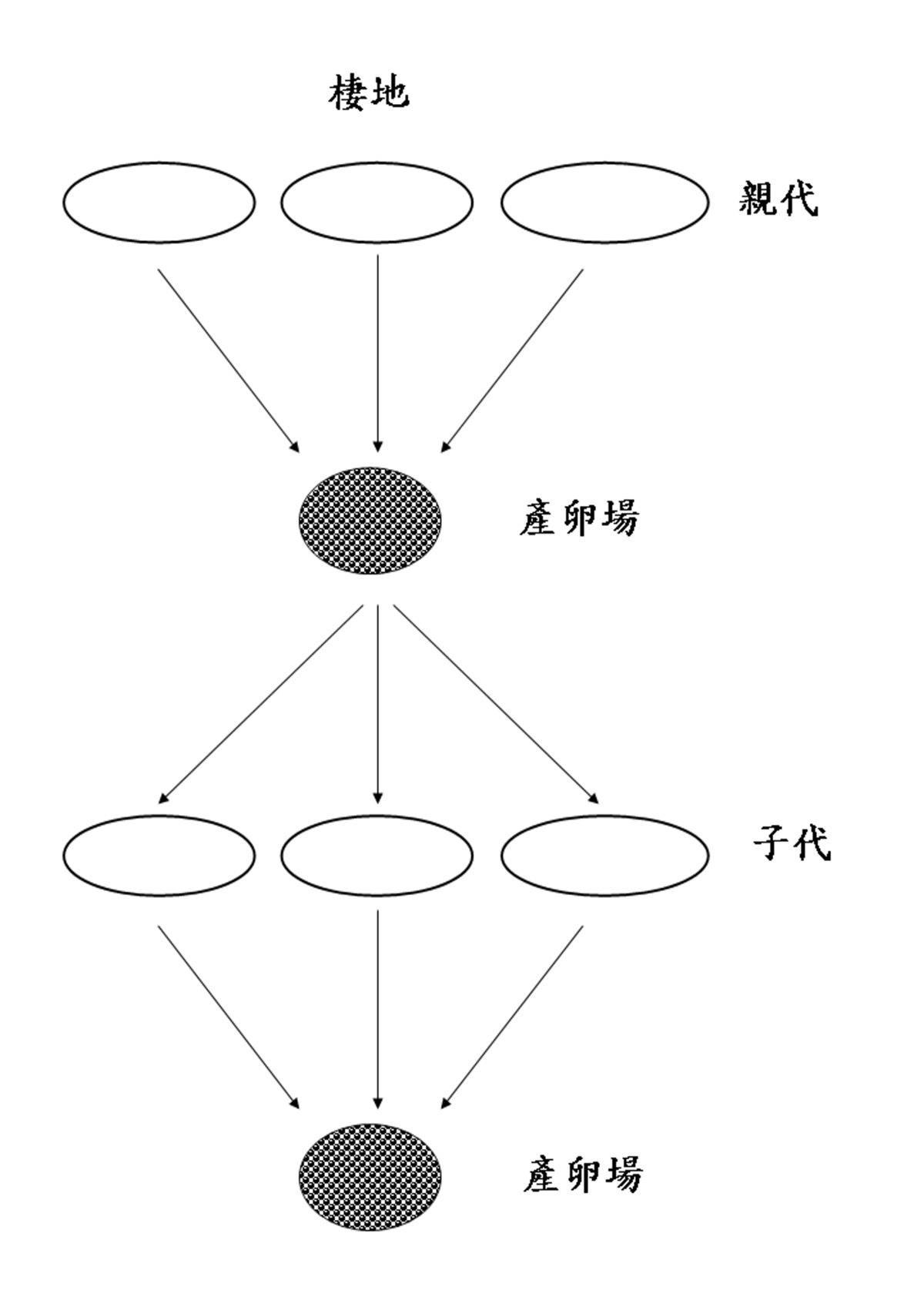
圖1.3.
20鰻魚的繁殖策略。每一種鰻魚都有一個固定的產卵場,不管子代擴散範圍有多廣,親魚最終都會回到出生的產卵場繁殖下一代。