鰻魚的生命從授精卵(Egg)開始、經過柳葉鰻(Leptocephalus)、玻璃鰻(Glass eel)、鰻線(Elvers)、黃鰻(Yellow eel)和銀鰻(Silver eel)等六個階段。最早科學家並不知道淡水鰻是在海裡產卵,只知道每年有大量的玻璃鰻來到河川的入海口,然後進入河川內長大成為黃鰻。就是看不到鰻魚產卵和牠生下的柳葉鰻。因此,引起科學家萬里尋跟的好奇心。
1.2.1 第一個揭開鰻魚生活史神秘面紗的科學家(The pioneer discovering the life history secret of eels)
約翰尼-史密特(Johannes Schmidt,1877-1933),是第一個揭開歐洲鰻生活史秘密的丹麥科學家(Schmidt 1922)。在約翰尼-史密特之前的研究者並不知道淡水鰻在海洋產卵,以及其生活史及體型有這麼曲折的變化,而把柳葉鰻當成不同的種類。經過契而不捨的追蹤,才發現丹麥境內的歐洲鰻是從美國佛羅里達東方海域的百慕達神祕三角洲附近的藻海(Sagasso Sea)誕生後,漂洋過海來到丹麥的(圖1.2.1)。這個發現引發日本人近60年來探索日本鰻產卵場的熱潮。

圖1.2.1(左圖)第一個揭開歐洲鰻生活史秘密的丹麥科學家約翰尼-史密特(Johannes Schmidt,1877-1933)。(右圖)歐洲鰻和美洲鰻在藻海(Sargasso Sea)產卵後,其柳葉鰻分別擴散到歐洲和美洲。紅星表示丹麥的歐洲鰻是從藻海來的
1.2.2日本鰻產卵場的探索歷史 (Historic exploration of the spawning ground of the Japanese eel)
日本科學家從1930年代起才開始調查太平洋的日本鰻產卵場。從日本近海開始順著黑潮一路往前追蹤,從柳葉鰻體型的愈採愈小,亦步亦趨而發現產卵場(圖1.2.2)。近60幾年來的調查,比較重要的發現分別是1968年在日本南方海域所捕獲體長60 mm的柳葉鰻,1975年田中昌一教授在台灣東方海域捕獲52尾體長50 mm左右的柳葉鰻(Tanaka 1975),1986年在菲律賓東方捕獲21尾體長40 mm柳葉鰻(Kajihara 1988),1988年和1989年在台灣東方捕獲體長20~30 mm的柳葉鰻(Ozawa
et al. 1989,1991)。最後是1991年在馬里亞納島西側採到1000多尾體長10 mm左右剛出生不久的前期柳葉鰻。經過60多年的探索,終於發現日本鰻的產卵場(Tsukamoto 1992)。
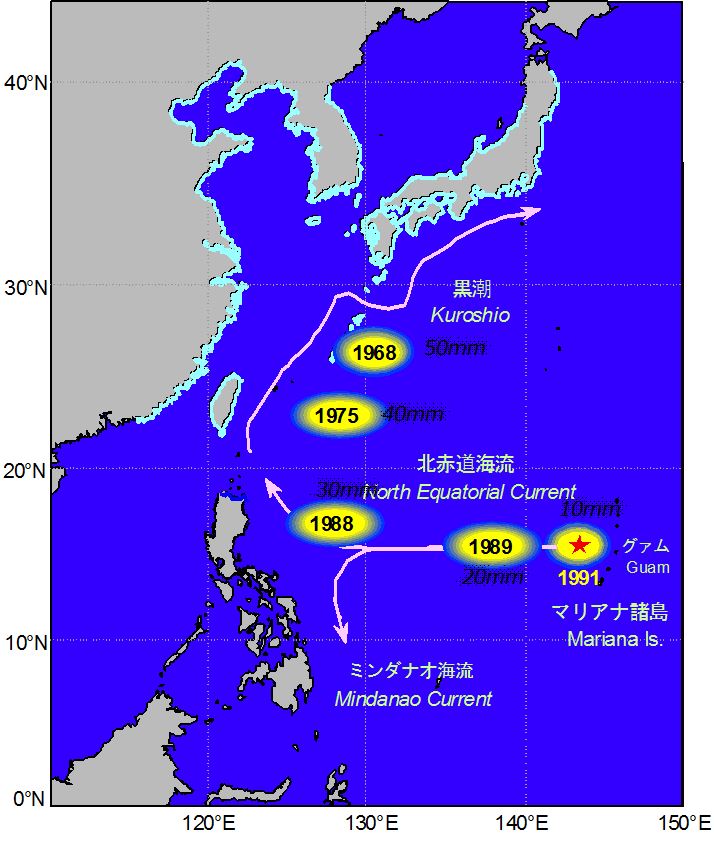
圖1.2.2日本研究團隊從愈採愈小的柳葉鰻體型,亦步亦趨而發現產卵場(photo: Katsumi Tsukamoto)
1.2.3產卵場發現的艱辛過程(Difficulties in discovering the spawning ground)
日本鰻產卵場的發現有如大海撈針,是一件艱辛的工作。產卵場的發現,有賴於地毯式地搜索柳葉鰻。日本東京大學海洋研究所(現在的大氣海洋研究所)研究船”白鳳丸” (圖1.2.3),跟據第5航次的密集調查,由柳葉鰻出現與否的位置研判,日本鰻產卵場應該在馬里亞納島西側,因為東側未出現柳葉鰻(圖1.2.4)。馬里亞納島以東是全世界最深的馬里亞納海溝,深度超過1萬2千公尺。

圖1.2.3日本東京大學海洋研究所探索日本鰻產卵場的研究船”白鳳丸”(photo: Katsumi Tsukamoto)
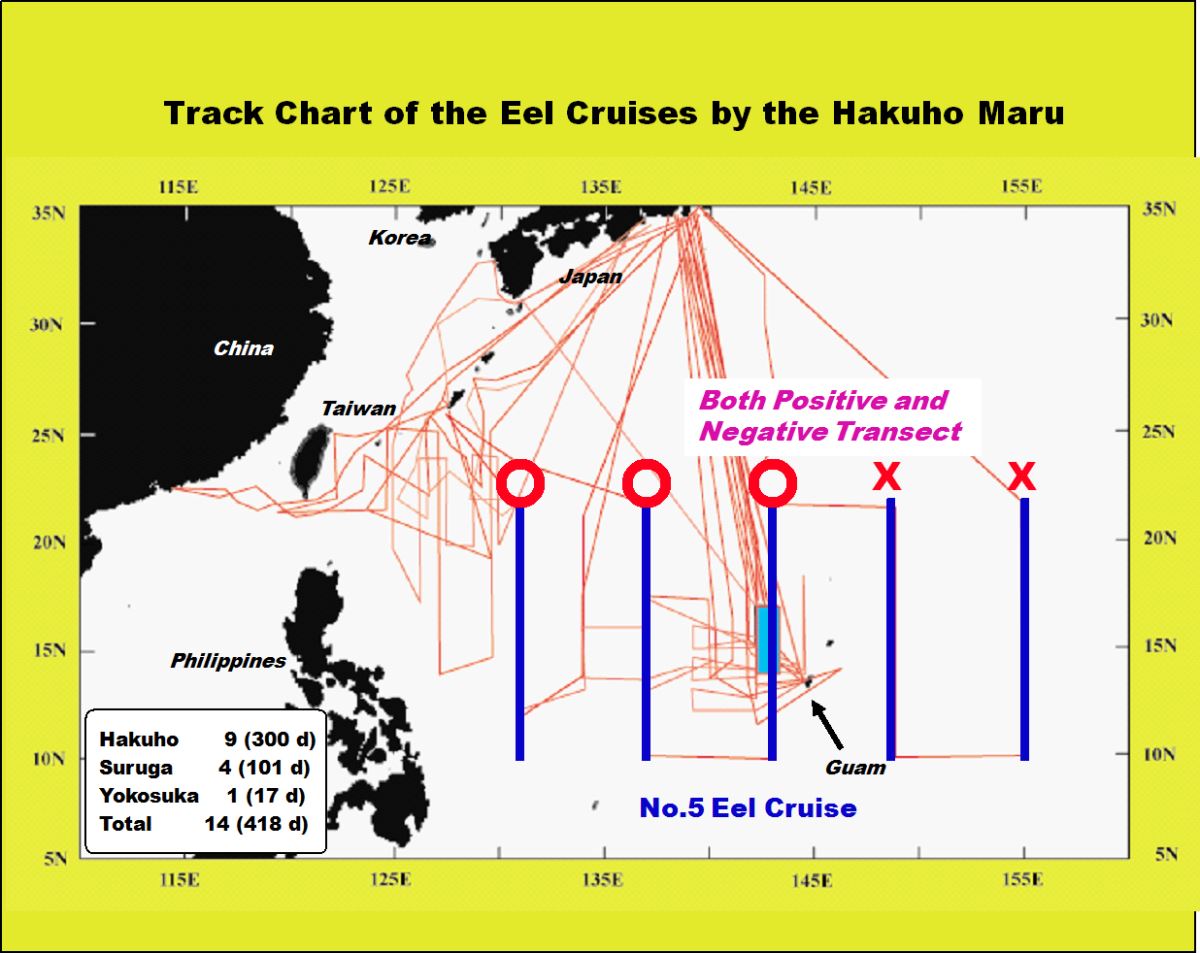
圖1.2.4研究船調查日本鰻產卵場的航跡圖和次數(天數)統計以及白鳳丸研究船第五次調查時柳葉鰻出現與否的航跡圖(Tsukamoto
et al. 2003,Eel Biology)
科學家到了產卵場海域後,先用大型浮游生物網採集浮游生物(圖1.2.5, 6),再從浮游生物中挑出鰻魚卵及柳葉鰻(圖1.2.7,8),最後再以DNA技術鑑定這些卵及柳葉鰻是否為日本鰻(圖1.2.9)。

圖1.2.5研究船”白鳳丸” 在馬里亞納島西側拖曳大型浮游生物網採集柳葉鰻的情景(photo: Katsumi Tsukamoto)

圖1.2.6採集柳葉鰻的大型浮游生物網(Kuroki& Tsukamoto 2012, Eel on the move)

圖1.2.7篩選大型浮游生物網採集的浮游生物(photo: Katsumi Tsukamoto)

圖1.2.8柳葉鰻及各種浮游生物(photo: Mari Kuroki)

圖1.2.9研究團隊利用DNA 技術鑑定海洋浮游生物中挑出的卵及前期柳葉鰻是否為鰻日本鰻(左起Dr. Aoyama, Dr. Otake, Prof. Tsukamoto, Dr. Kuroki and Dr. Sudo)(photo: Katsumi Tsukamoto)
1.2.4有助於日本鰻產卵場發現的兩個假說(Two hypotheses helpful in discovering the spawning ground of the Japanese eel)
日本鰻產卵場的發現,看似簡單,卻是花很漫長的時間、累積不少錯誤的經驗才成功的。日本鰻在陸地河川分布的最南限是台灣。因此,研究鰻魚的泰斗松井魁博士(1957),認為日本鰻產卵場應該在台灣東部的沖繩海溝附近,這是一個錯誤的假說,也誤導了日本鰻產卵場調查的方向(Kimura
et al. 1994)。東京大學的田中昌一教授為了驗證日本鰻的產卵場是否位於台灣東方海域,1972年在台灣東部進行柳葉鰻的採集調查,但只採獲大型的柳葉鰻,離產卵場還有一段距離。
台灣水產試驗所郭河先生(1971)也曾經提出日本鰻的產卵場假說,他想如果松井博士所言屬實,日本鰻果真在台灣東部海域產卵的話,那麼生下來的玻璃鰻不就全部被黑潮輸送到台灣以北的中國大陸、韓國及日本地區了嗎? 那台灣西海岸的鰻苗從那裡來?於是郭先生認為台灣西南部琉球海溝附近應該有另一個產卵場,否則無法解釋台灣西部河口域鰻苗的出現(圖1.3.14)。這個錯誤的假說,也讓台灣水產試驗所,從1976年到2003年為止,一直在小琉球附近進行種鰻放流。產卵場的錯誤判斷,是因為不瞭解鰻魚的生活史、仔魚輸送過程及海流構造所致。
過去認為鰻魚在冬季產卵。到了1980年之後,因為耳石日周輪的研究(Tabeta
et al. 1987),逐漸對日本鰻魚的產卵季節有了進一步的認識。原來每年冬季(12月~1月)在台灣沿岸大量出現的鰻線,是半年前的夏天(5~6月)誕生的(Tsukamoto 1990, Tzeng 1990, Cheng and Tzeng 1996, Tsukamoto
et al. 2003)。由鰻魚誕生到鰻線被捕的半年海上漂流時間和海流的速度推算,日本鰻的產卵場應該離台灣很遠,要往黑潮的源頭(北赤道洋流)推進才合理(圖1.2.2)。於是東京大學Kajihara教授在1986年8月的退休航海時,在菲律賓東方海域利用IKMT中層拖網進行柳葉鰻的採集調查,採獲了一批體型30~40mm的柳葉鰻,筆者也有幸參與此次白鳳丸(KH-86-4)航次的鰻魚產卵場調查(Tzeng 1988)。同時登船者,還有接受IOC/UNESCO 補助,來自泰國的Dr. R. Chayakul 和來自韓國的李泰源博士(圖1.2.10)。從此次鰻魚產卵場調查採集的柳葉鰻體型大小來看,推測調查區離產卵場已經不遠了,因而奠定了日後尋找日本鰻產卵場的正確方向 (Kajihara
et al. 1988)。

圖1.2.10 參與白鳳丸(KH-86-4)航次的鰻魚產卵場調查(左起筆者、Kajihara教授、Dr. R. Chayakul 和李泰源博士)
不出所料,日本東京大學的研究團隊於1991年日本鰻產卵場的密集調查中,果然在馬里亞納島西側採獲了1000多尾剛出生不久的前期柳葉鰻(Tsukamoto 1992)。這個結果讓東京大學的研究團隊非常興奮,因為日本鰻產卵場的位置就近在咫尺。
後來,日本水產廳利用開洋丸調查船的大型拖網(圖1.2.11),更進一步於2008年3月和2008年6月分別採獲日本鰻的銀鰻(圖1.2.12)及其受精卵,日本鰻產卵場的位置終於得到驗證。
日本鰻產卵場的精確位置之發現,有賴以下兩個假說:

圖1.2.11日本水產廳利用開洋丸的大型拖網在產卵場採集日本鰻銀鰻(photo: Hiroaki Kurogi; copyright: Fisheries Agency, Japan)

圖1.2.12日本水產廳的研究人員於2008年6月3日在馬里亞納島西側海山的日本鰻產卵場採獲的銀鰻(photo: Seinen Chow, copyright: Fisheries Agency, Japan)
(1)新月假說(New moon hypothesis)
能夠精準掌握日本鰻產卵場的位置,要歸功於兩個重要的理論基礎。其一,是新月假說(Tsukamoto
et al. 2003,Tsukamoto 2006)。因為新月假說才得以正確掌握鰻魚的產卵時間,採獲日本鰻的受精卵和柳葉鰻來證明產卵場位置。因此,1991年7月日本東京大學白鳳丸才能成功地在馬里亞納島西側採穫1000多尾柳葉鰻(Tsukamoto 1992)。這些柳葉鰻的體長分布如圖1.2.13所示。如果把每一尾柳葉鰻的耳石取出,計算其日周輪(圖1.2.14),便可推算圖1.2.13的1000多尾柳葉鰻的出生日期(圖1.2.15)。有趣的是這些柳葉鰻的出生日都集中在5月的新月當天,或6月新月後1~3天(Tsukamoto et al. 2003)。有了這個訊息就可預期母鰻產卵的時間,而找到日本鰻的正確產卵場。
柳葉鰻的耳石日周輪,在電子顯微鏡下可看到中心直徑約10µm的黑色深凹部份,這是鰻魚孵化之前的胚胎期,之後從孵化輪到第一個攝食輪的放射狀的結晶部分,為卵黃囊期,第一輪之後為前期柳葉鰻開口攝餌之後所形成的日周輪(圖1.2.14)。前期柳葉鰻大約5天後開始攝餌,之後隨著日夜周期的變化,每天形成一輪。因此,由日周輪數加5天,就是柳葉鰻的實際日齡。換言之,由日周輪數就可以知道鰻魚的出生日。
1.2.13、柳葉鰻體長分布(photo:Tsukamoto)

圖1.2.14、柳葉鰻的耳石日周輪。圖中紅點分別為孵化輪和第一次攝食輪,之後每天形成一輪。由總輪數及柳葉鰻被捕獲的日期就可回推柳葉鰻的出生日(photo: Mari Kuroki)
圖1.2.15、柳葉鰻出生日的頻度分佈(photo: Katsumi Tsukamoto)
(2)海山假說(Sea mount hypothesis)
海山假說是發現日本鰻產卵場的第2個重要關鍵。大海茫茫,要捕獲正在產卵的母鰻或其授精卵來證明產卵場,談何容易。海山是海洋中接近水面的島嶼,山的背流面水流比較暖和,而且有隱蔽物,可能是雌雄鰻魚約會交配繁殖下一代的理想地點。將目前為止採獲日本鰻的卵、前期柳葉鰻及產卵親魚的地點,描繪到海底地形圖上,則發現日本鰻的產卵場集中在西馬里亞納海嶺的南端西側(圖1.2.16,17)(Tsukamoto et al. 2011),稍往南就是全世界最深的馬里亞納海溝的挑戰者深淵。
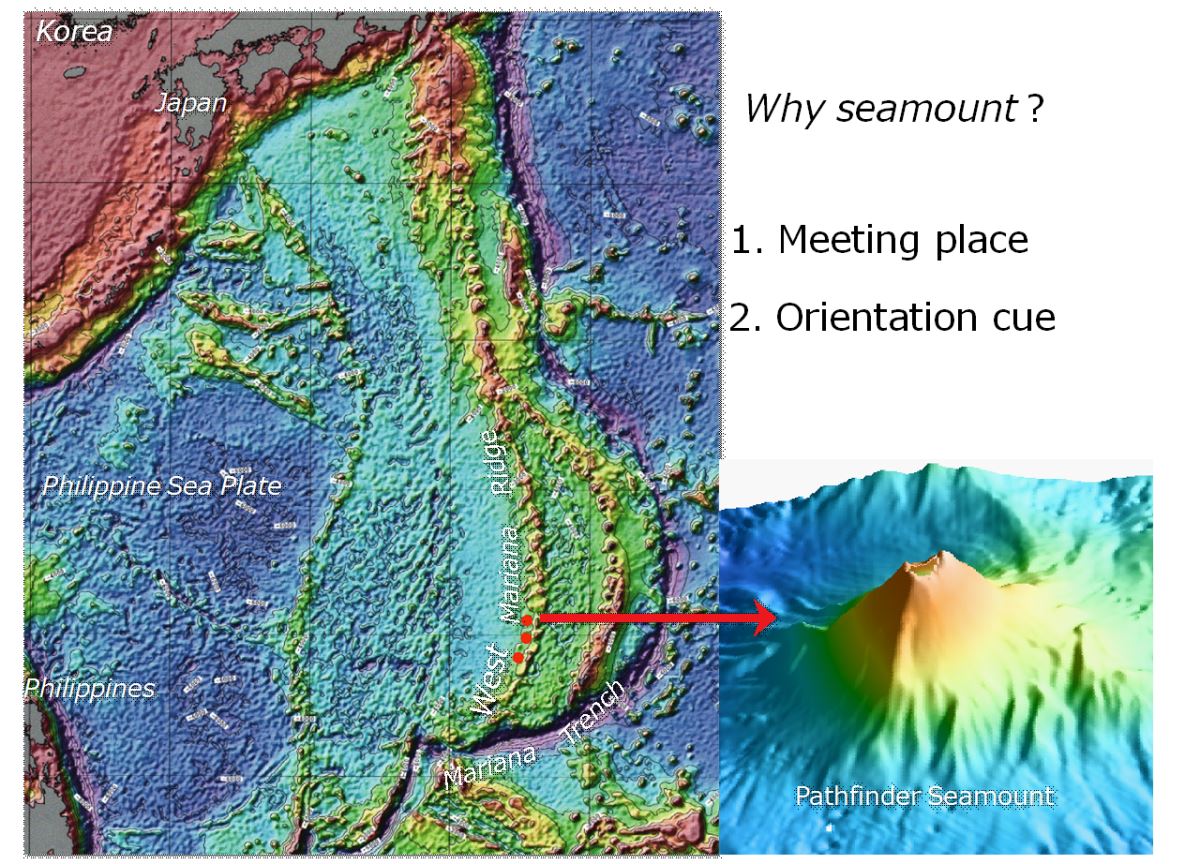
圖1.2.16日本鰻的產卵場位於西馬里亞納海嶺南端的海山(photo: Kyoko Okino, The University of Tokyo & JAMSTEC)
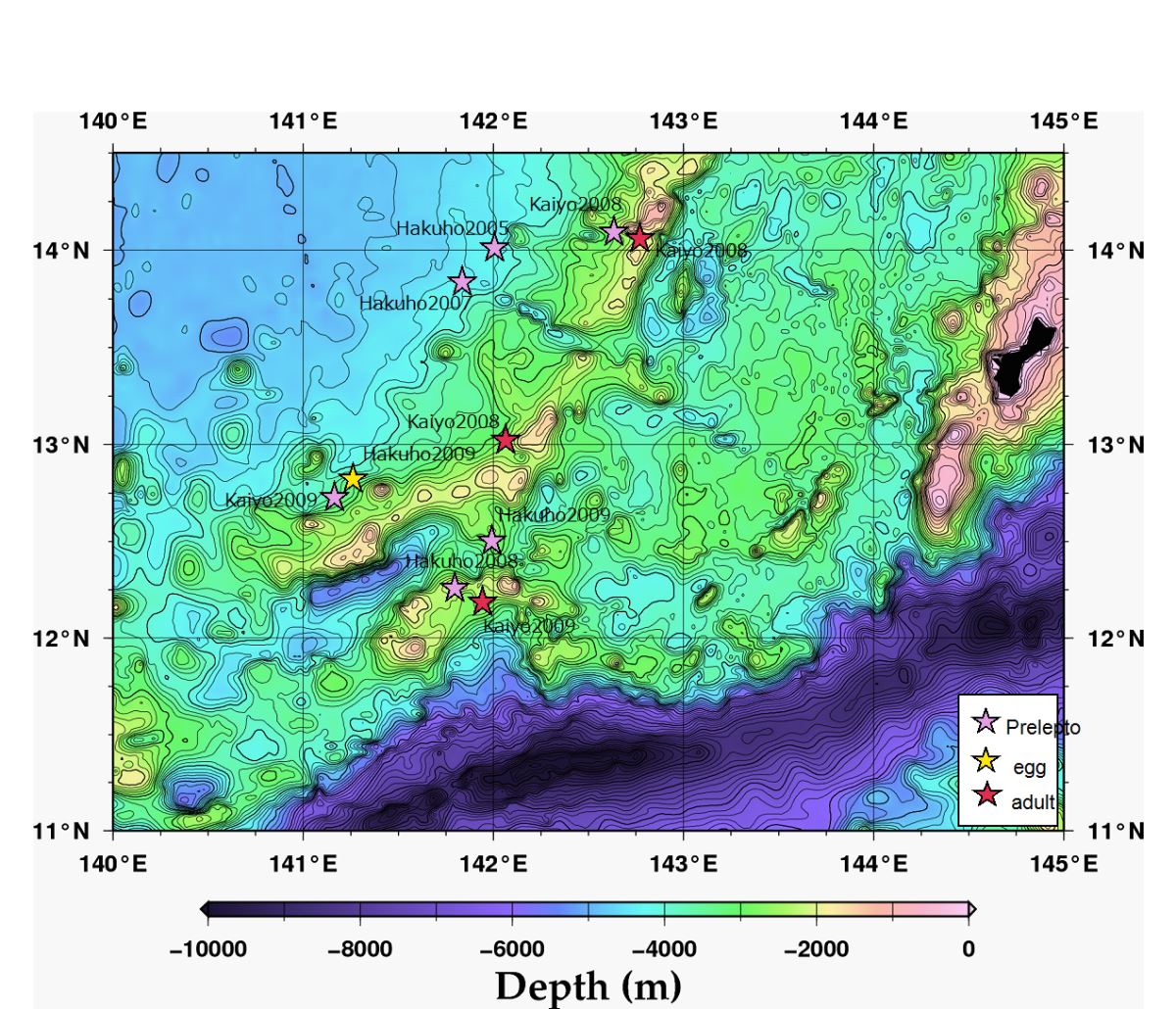
圖1.2.17在西馬里亞納海嶺南端的海山採獲日本鰻親魚及其生下的卵和前期柳葉鰻(photo: Hiroaki Kurogi and Katsumi Tsukamoto)
鰻魚為何在西馬里亞納海嶺產卵,仍然是個謎。這個地方就是菲律賓板塊的東南角,大約3-4千萬年前,地球板塊開始分裂時日本鰻不知受什麼力量的指引來到此地產卵也說不定。分布在東北亞的日本鰻從產卵場出發,要游3,000 km以上才能到達陸地河川成長。
1.2.5 台灣學者第一次採獲柳葉鰻(The first collection of leptocephali by Taiwanese scientists)
1995年8月16日至23日,農委會水產試驗所派出水試一號試驗船(圖1.2.17),於 12
o30'N - 14
o30'N與 131
o30'E - 140
o30'E之間,也就是菲律賓以東的海域,捕獲 27 尾柳葉鰻。其中三尾是日本鰻。這是台灣有史以來,第一次捕獲日本鰻的柳葉鰻,其意義非凡(圖1.2.18)。三尾柳葉鰻的體長分別為 27.4 mm、28.8 mm 以及31.4 mm。由耳石日周輪數目研判,其日齡分別為46天、50天及51天。孵化日期為7月1日及3日。相當於農曆的6月2日及4日的新月期間(Liao
et al. 1996)。此結果也驗證了鰻魚產卵期的新月假說。

圖1.2.18台灣行政院農業委員會水產試驗所及水試一號試驗船

圖1.2.19、水試一號試驗船於1995年8月16日至23日在菲律賓以東的海域捕獲三尾日本鰻的柳葉鰻(黑色圓圈)。